 Original Article
Original Article

Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
†These authors shared the first authorship and contributed equally to this work.
ORCID: https://orcid.org/0000-0001-7588-3846
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
†These authors shared the first authorship and contributed equally to this work.
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
ORCID: https://orcid.org/0000-0003-4457-1422
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
ORCID: https://orcid.org/0000-0003-4789-661X
Affiliation:
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
Affiliation:
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
ORCID: https://orcid.org/0000-0001-8314-8091
Affiliation:
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
Affiliation:
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
ORCID: https://orcid.org/0000-0002-2518-2803
Affiliation:
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
ORCID: https://orcid.org/0000-0003-0068-2614
Affiliation:
1Centro de Investigación en Ciencias de la Salud (CICSA), FCS, Universidad Anáhuac México Campus Norte, Huixquilucan, Mex. 52786, México
2Escuela Militar de Graduados de Sanidad, Centro de Investigación y Desarrollo del Ejército y Fuerza Aérea Mexicanos, Universidad del Ejército y Fuerza Aérea, México City 11200, México
Email: jose.ibarra@anahuac.mx
ORCID: https://orcid.org/0000-0003-2489-4689
Explor Neuroprot Ther. 2026;6:1004160 DOI: https://doi.org/10.37349/ent.2026.1004160
Received: August 11, 2025 Accepted: April 24, 2026 Published: June 25, 2026
Academic Editor: Rodrigo Pacheco, Universidad San Sebastián, Chile
The article belongs to the special issue Role of Microbiota in Neurological Diseases
Aim: Spinal cord injury (SCI) disrupts autonomic regulation of the gastrointestinal tract, resulting in altered motility, microbial dysbiosis, and decreased production of short-chain fatty acids such as butyrate, a metabolite with neuroprotective properties. This study aimed to evaluate the effects of SCI level and severity on gut microbiota composition and butyrate concentration, and to determine the impact of symbiotic supplementation with Enterococcus faecium and agave inulin on microbial and metabolic recovery.
Methods: Twenty female Sprague Dawley rats were randomized into five groups: Sham, thoracic five moderate (T5M), thoracic five severe (T5S), thoracic nine moderate (T9M), and thoracic nine severe (T9S). Fecal samples were collected four weeks post-injury and again after four weeks of daily symbiotic administration. Butyrate levels were quantified by gas chromatography, and microbial composition was analyzed by 16S ribosomal RNA (16S rRNA) sequencing (V3–V4 region). Locomotor recovery was assessed weekly using the Basso, Beattie, and Bresnahan (BBB) scale.
Results: All SCI groups displayed significant dysbiosis compared with Sham, characterized by decreased relative abundance of butyrate-producing genera such as Clostridium at week four, as confirmed in the main dataset. Following symbiotic supplementation, Bifidobacterium, Lactobacillus, and Clostridium levels showed partial restoration, although responses varied by injury site and severity. The T9M group demonstrated the most consistent recovery of butyrate-producing taxa; however, butyrate concentration decreased after treatment, while T9S showed a significant increase in butyrate concentration, and T5S exhibited minimal change. Motor recovery was greatest in T9M, followed by T9S and T5M, with no significant improvement in T5S.
Conclusions: SCI level and severity strongly influenced microbial dysbiosis, butyrate metabolism, and motor recovery. Symbiotic supplementation partially restored butyrate-producing bacterial taxa, with the most pronounced metabolic and functional benefits observed in the T9M group. These findings support a level-dependent role of gut-brain axis modulation as a potential therapeutic strategy after SCI.
Gut microbiota, particularly that located in the cecum and colon, express carbohydrate-activated enzymes, allowing them to produce complex carbohydrates through anaerobic fermentation, generating metabolites such as short-chain fatty acids (SCFAs) [1] which work as pleiotropic signaling compounds [2] in the microbiota-gut-brain/spinal cord axis. Briefly, in this axis, signals from the brain influence motor, sensory, and secretory function of the gut, while visceral messages from the gut impact brain function [3], constituting a continuous feedback loop.
The gastrointestinal tract (GIT) serves as the main interface for interaction and nutritional exchange between the animal and the external environment [4]. Bacterial composition can be influenced by several factors, including immune mechanisms, diet, and gut motility, and stressors such as sepsis, burns, trauma, and infection [5, 6].
Spinal cord injury (SCI) affects sensory, motor, and autonomic functions, exerting significant impacts on the physical and psychosocial well-being of patients [7, 8]. Complications following SCI depend directly on the level, the intensity, and the type of injury. Symptoms can be classified by severity based on the neurological level: cervical, thoracic, lumbar, or sacral. For instance, patients with damage above the T5 level require additional care to prevent comorbidities related to bowel dysfunction (BD) or neurogenic bladder. At this level, supraspinal sympathetic pathways to the splanchnic and pelvic nerves are disrupted, impairing autonomic control of intestinal motility and urinary sphincter tone. This autonomic imbalance contributes to chronic constipation, fecal impaction, and urinary retention, which are common and clinically significant complications in high-thoracic injuries [7, 9].
BD is one of the most common complications after traumatic SCI, affecting 25–41% of all SCI cases [10–12]. BD involves changes in the extrinsic autonomic innervation to the bowel, with clinical repercussions in motility (i.e., constipation in 46%, fecal incontinence in 41%), sphincter control (31%), and abdominal cramping (18%) [12, 13]. While intrinsic enteric innervation remains intact, it may lose its integrity over time due to alterations in the extrinsic system [14].
This series of events leads to gut dysbiosis, which has been documented by various authors in both patients and animals [15–17]. Some studies have explored therapeutics aimed at re-establishing these alterations, such as nutraceuticals or sodium butyrate administration [16, 18]. The role of Enterococcus faecium as a nutraceutical has been investigated, demonstrating immunomodulatory effects in various pathologies [19]. It has been shown to increase transforming growth factor-beta (TGF-β) expression in the GIT during infectious disorders [20].
The E. faecium + agave inulin combination was selected due to its complementary mechanisms targeting both microbial restoration and host immune modulation. E. faecium exhibits strong mucosal adhesion, acid and bile tolerance, and proven efficacy in restoring gut eubiosis after inflammatory insults. It enhances intestinal TGF-β expression, reduces IL-6 and TNF-α release, and modulates toll-like receptor signaling, collectively supporting anti-inflammatory and barrier-protective effects [19, 20]. Agave inulin, a highly fermentable fructan, selectively stimulates the growth of beneficial genera such as Bifidobacterium and Lactobacillus, promoting the synthesis of SCFAs including acetate and butyrate [21, 22]. The synergistic action of this symbiotic pair has been reported to improve gut permeability, immune tolerance, and microbial diversity in models of gastrointestinal inflammation [23, 24]. Therefore, this formulation was considered mechanistically suitable to counteract SCI-induced dysbiosis and to enhance butyrate recovery through both microbial and host-mediated pathways.
One reported combination therapy involved agave inulin and E. faecium; both in vitro and clinical studies have indicated that agave inulin promotes the growth of Bifidobacterium, enhancing lactic and acetic acid production while reducing pathogenic bacterial strains [23, 24]. On the other hand, E. faecium has effects on both innate and adaptive responses of the immune system and can modulate pro-inflammatory cytokines, evidenced by its ability to reduce the expression of IL-8 and IL-10 while increasing TGF-β expression in the GIT mucosa during infectious processes [23, 24].
The main goal was to characterize gut microbiota and fecal butyrate concentrations in rats with different intensities and levels of SCI before and after a symbiotic administration using E. faecium and agave inulin.
In addition, identifying correlations between changes in gut microbiota composition, fecal butyrate levels, and motor recovery may help to elucidate the functional interplay between microbial metabolism and neurophysiological outcomes following SCI, providing a foundation for future mechanistic studies.
SCI and all experimental procedures were conducted in accordance with protocols approved by the National Institutes of Health (US) guidelines and the Guide for the Care and Use of Animals. The Animal Bioethics and Welfare Committee of the Universidad Anáhuac México approved all animal procedures (Registry number: 201702-CI) in accordance with the National Institutes of Health Guide for the care and use of laboratory animals, and the Mexican Official Norm on Principles of Laboratory Animal Care (NOM 062-ZOO-1999).
Twenty female Sprague-Dawley rats, aged eight to eleven weeks old, weighing between 200 g and 300 g, were housed into five groups of four rats each (n = 4/group) under a 12-hour light/dark cycle, in thermally controlled rooms. They were fed standard pellets and given water ad libitum. To minimize stress, all animals were handled at least once daily for 30 days prior to surgery, and environmental enrichment was provided. Sterile bedding and filtered water were replaced daily. Females were selected to minimize variability in body weight, stress-related autonomic responses, and post-injury urinary complications, which are more frequent in males and could confound gastrointestinal outcomes and fecal sampling. This approach also ensured consistency with prior SCI studies from our group that employed the same sex and strain, allowing for valid comparisons across experiments. The symbiont was administered daily by orogastric tube for four consecutive weeks (weeks 4–8 post-injury). To minimize potential stress or motility alterations due to the orogastric procedure, animals were gently handled and acclimated for three days before administration, and no signs of distress or reduced food intake were observed.
The rats were allocated into five groups using the STATS TM v.2.0 software: 1) Sham (n = 4), 2) severe injury at T5 level (thoracic five severe, T5S) (n = 4), 3) moderate injury at T5 level (thoracic five moderate, T5M) (n = 4), 4) severe injury at T9 level (thoracic nine severe, T9S) (n = 4), and 5) moderate injury at T9 level (thoracic nine moderate, T9M) (n = 4). The five groups were studied over a period of eight weeks. Four weeks post-injury (wpi), all groups received a symbiotic consisting of E. faecium and inulin. Stool samples were collected at 4 wpi (prior to symbiotic administration) and at 8 wpi (after four weeks of symbiotic treatment). Gut microbiota composition and butyrate concentrations were analyzed from fecal samples at both time points. Motor recovery was assessed weekly in all groups until the end of the experiment (8 wpi). The sample size calculation for studies evaluating the taxonomic composition of the gut microbiota was performed using the formula proposed by Scheff et al. [25], 2003 which estimates power based on pairwise distance matrix comparisons between groups. This calculation indicated that four rats per group would provide 90% power to detect significant differences in microbial composition between pre- and post-treatment conditions, taking into account both injury level and severity.
The study followed a pre- and post-test longitudinal design in which each rat served as its own control. Fecal samples collected at 4 wpi (pre-treatment) and 8 wpi (post-treatment) were compared within each group to assess the effect of the symbiotic. This approach minimized inter-animal variability and reduced total animal use in accordance with ethical principles.
Rats were weighed and anesthetized with ketamine (90 mg/kg) and xylazine (10 mg/kg) administered intraperitoneally. After fifteen minutes, a laminectomy was performed at the thoracic vertebra to expose the dura mater. Stabilization clamps were used to access the T5 or T9 segment of the spinal cord. Injuries were inflicted using the Infinite Horizons SCI device 5.0, programmed for moderate injury at 200 kilovolts and severe at 300 kilovolts.
Post-surgery, all rats received 10 mg/kg of paracetamol orally twice a day and 5 mg/kg of ketoprofen, once a day via subcutaneous injection for one week, along with fluid replenishment with saline solution. Manual emptying of the bladder and rectum was performed three times daily until sphincter control was recovered. Sham-operated animals underwent all surgical procedures to expose the spinal cord but were not injured and served as controls. Animals were carefully monitored for signs of infection, dehydration, or self-mutilation, with appropriate veterinary care provided as needed. All experiments were designed and reported in accordance with established guidelines. At the end of the experiment (8 wpi), for euthanasia, the animals were first anesthetized intraperitoneally, with an injection of a ketamine (90 mg/kg) and xylazine (10 mg/kg) mixture. Once the animals no longer exhibited reflexes, an intraperitoneal injection of sodium pentobarbital (200 mg/kg, lethal dose) was administered.
Treatment began four wpi, rats were weighed every third day, and a homogeneous mixture of E. faecium (0.8 g equivalent to 1.6 × 1010 CFU/kg body weight) plus agave inulin (0.86 g/kg body weight) was administered via an orogastric tube, diluted in drinking water until the end of the experiment (8 wpi). The symbiotic consisted of E. faecium [1 × 107 CFU; Alimentos Esenciales para la Humanidad (AEH) S.A. de C.V., Ciudad de México, México] and agave inulin (8 g; AEH S.A. de C.V., Ciudad de México, México).
Fecal samples were collected individually from each rat at 4 and 8 wpi using sterile forceps in a laminar flow hood. Each sample was stored at –80°C until processing. For microbial characterization, genomic DNA was first extracted individually from 0.25 g of feces using the QIAamp PowerFecal DNA Kit (QIAGEN, 11993). To obtain representative community profiles for each experimental group and time point, equal amounts of purified DNA from each individual within the same group were pooled prior to 16S ribosomal RNA (16S rRNA) gene amplification and sequencing on the Illumina MiSeq platform. This approach was chosen to ensure sufficient DNA yield and sequencing depth while reducing technical variability between samples. Consequently, microbial composition data reflect group-averaged taxonomic profiles, whereas metabolic (butyrate) and behavioral analyses were performed on individual samples. For sequencing, equal volumes of DNA extracted from each animal of the same experimental group and time point were pooled to obtain one composite sample per group. The pooled DNA was subsequently divided into four technical aliquots, each processed independently through the 16S V3–V4 library preparation (Nextera XT kit, Illumina) and sequenced on the MiSeq platform.
Stool samples were collected at 4 wpi (before symbiotic administration) and at 8 wpi (after four weeks of symbiotic administration) through manual extraction in a disinfected laminar flow hood. After handwashing and donning gloves, gentle pressure was applied to the rectal ampulla, and samples were collected with sterile Adson tissue forceps, previously sterilized with 12% Krit germicidal solution. Samples were stored in 2 mL centrifuge tubes at –80°C for further processing.
To isolate and purify bacterial DNA, 0.25 g of feces was processed using the QIAamp PowerFecal DNA Kit (QIAGEN Model: 11993), following the manufacturer’s protocol. Sample quality was assessed using the Thermo Scientific NanoDrop One Microvolume UV-Vis Spectrophotometer, measuring at a wavelength range 260–320 nm to obtain quantity (ng/mL) and assess purity using the ratios 260/280 nm and 260/320 nm (values above 1.8 were included). Electrophoresis (1% agarose gel) was performed to evaluate DNA integrity.
Sequencing was performed on MiSeq following the workflow for preparing 16S rRNA Gene Amplicons for the Illumina MiSeq System (Part # 15044223 Rev. B): amplicon primers, library preparation, and sequencing on MiSeq.
For the 16S library preparation, modifications were made. The conditions for the first stage PCR were 98°C for 30 seconds, followed by 5 cycles of 98°C for 20 seconds, 55°C for 30 seconds, and 72°C for 30 seconds, after which a final elongation step at 72°C for 5 minutes, and then held at 4°C. The second stage PCR involved 98°C for 30 seconds, followed by 8 cycles of 98°C for 20 seconds, 55°C for 30 seconds, and 72°C for 30 seconds, concluding with a final elongation step at 72°C for 5 minutes, and then held at 4°C.
Once genetic material was obtained, PCR was performed, and sequences were purified. Operational Taxonomic Units (OTUs) were identified. For classification analysis, we used the 16S Metagenomics software, available on the BaseSpace platform (Illumina, Inc.), utilizing a modified version of the Greengenes taxonomic database. The following metrics were reported for each sample: 1) alpha diversity: using the Shannon index and the number of identified species (R), and 2) beta diversity.
Chimeric sequences were discarded, and all non-chimeric input sequences were required to match at least one OTU with over 97% identity. Dissimilarity scores were calculated using the Bray-Curtis distance metric, considering relative abundance at the genus level for each rat and lesion group. Hierarchical clustering and principal coordinate analysis (PCoA) were employed to graphically summarize inter-sample relationships, with a heat map representing the relative abundance of the eight principal genera before and after symbiotic administration.
Fecal samples were collected by groups 4 wpi and 8 wpi. Prior to collection, sterile bedding was changed, and samples were harvested after an 8-hour waiting period using sterile tissue forceps. Pooled samples were stored in 50 mL centrifuge tubes at –80°C until processing.
Stool was lyophilized at –65°C under 1.33 × 10–6 torrs. One gram was weighed into a 50 mL centrifuge tube and 5 mL of distilled, deionized water was added. The mixture was homogenized for 10 minutes at maximum speed in a vortex mixer. Subsequently, the pH was evaluated with a pH meter, and it was acidified with 5 M HCl to the range of 2 to 3. The sample was centrifuged at 4,000 rpm for 25 minutes at 16°C, and 2 mL of chloroform was added. After a second centrifugation at 4,000 rpm for 2 minutes at 16°C, the aqueous phase was transferred to sterile 50 mL centrifuge tubes.
Butyric acid was determined by gas chromatography (GC) with a Shimadzu GC2010 (Japan), equipped with a silica capillary column for free fatty acids (model DB-1701, 30 m length, 0.25 mm internal diameter and 0.25 µm thickness, Agilent, USA). GC conditions included helium as carrier gas with a flow of 4 mL/min. The oven temperature was set to 250°C, with the column maintained at 95°C. The flow rates for hydrogen and air were set at 40 and 400 mL/min, respectively. The flame ionization detector (FID) was operated at 300°C. The retention time for butyric acid was between 3.5 and 3.6 minutes, and the injected sample volume was 2 µL, with butyric acid concentration at 0.1% w/v and the molecular weight of 88.11 g/mol. Each analysis lasted 20 minutes, with butyrate levels determined using Sigma-Aldrich standard 107-92-6. For each pooled sample, butyrate concentration was reported in µg butyric acid/2 µL. All the pools were processed in triplicate.
To evaluate motor recovery, animals were examined weekly by blinded raters using the Basso, Beattie, and Bresnahan (BBB) locomotor rating scale [25, 26] from the time of surgery until the end of the study.
Statistical analyses were conducted using GraphPad Prism v9.0 and PAST v.4.11. Normality was evaluated using the Shapiro-Wilk test. Since most data (e.g., BBB scores, microbial diversity indices) did not follow a normal distribution, non-parametric analyses were performed. For alpha-diversity indices [Richness (R) and Shannon; Figure 1], comparisons between groups were analyzed by the Kruskal-Wallis test, and pre- vs. post-treatment comparisons within each group by the Friedman test. Beta-diversity analysis (Figure 2) was performed using Bray-Curtis dissimilarity matrices and analyzed with PERMANOVA (Adonis) to estimate the explained variation (EV, EV%) between groups. Butyrate concentrations were compared between groups using the Kruskal-Wallis test, and pre- vs. post-treatment differences were evaluated with the Wilcoxon signed-rank test. Locomotor performance (BBB scores) was reported as median ± interquartile range (IQR). Longitudinal analysis of BBB recovery was performed using the Friedman test, and between-group comparisons at final time points by the Kruskal-Wallis test. All analyses were two-tailed, and p < 0.05 was considered statistically significant.
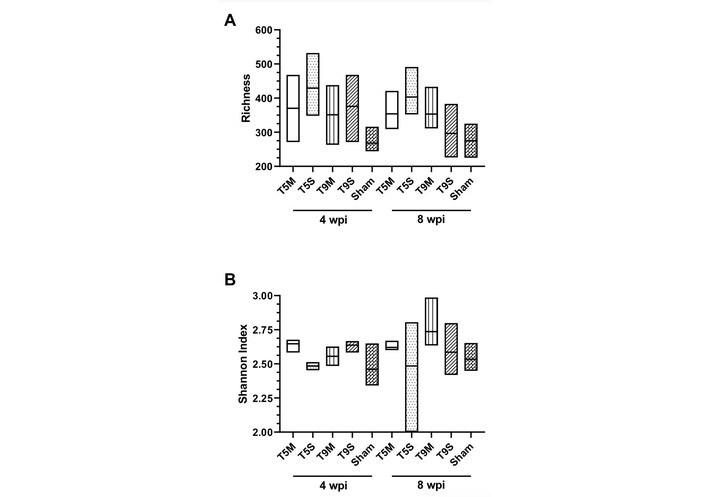
Comparison of means of Richness (R) and Shannon index before (4 wpi) and after (8 wpi) symbiotic administration. Box plots showing R (A) and Shannon index (B) across experimental groups (T5M, T5S, T9M, T9S, Sham) at 4 wpi (before symbiotic administration) and 8 wpi (after symbiotic administration). wpi: weeks post-injury; T5M: thoracic five moderate; T5S: thoracic five severe; T9M: thoracic nine moderate; T9S: thoracic nine severe.
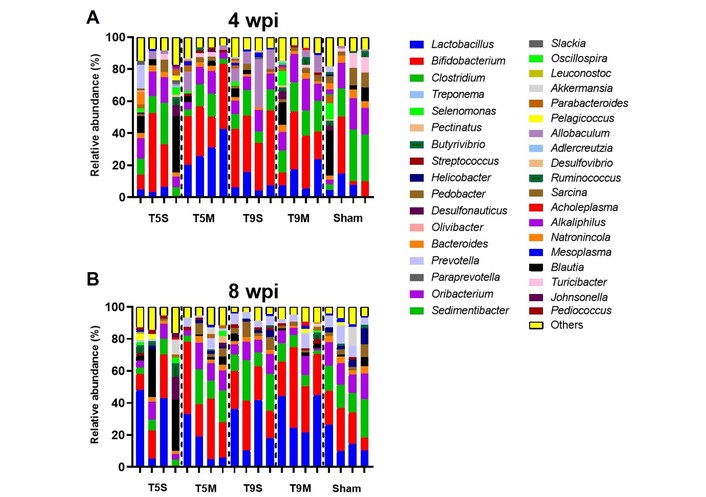
Relative abundance analysis of the gut microbiome (Beta diversity) before and after symbiotic administration. Agglomerative hierarchical clustering highlights the OTUs with the most significant differences in abundance between groups. Stacked bar plots showing the relative abundance of bacterial taxa across experimental groups (T5M, T5S, T9M, T9S, Sham) at 4 wpi (A—before symbiotic administration) and 8 wpi (B—after symbiotic administration). Each bar represents a technical replicate derived from pooled DNA samples of all animals within each group and time point. Colors indicate different bacterial taxa, and bar length corresponds to relative abundance. Following symbiotic supplementation, microbiome composition showed a trend toward homogenization across groups, with increased representation of beneficial genera such as Bifidobacterium and Lactobacillus. Enterobacteriaceae is presented as a family-level taxon (not genus-level) and is labeled accordingly in the figure. OTUs: Operational Taxonomic Units; wpi: weeks post-injury; T5M: thoracic five moderate; T5S: thoracic five severe; T9M: thoracic nine moderate; T9S: thoracic nine severe.
A total of 40 samples were sequenced. The characterization of the microbiota was performed on individual samples from each lesion group, both before and after symbiotic administration (4 wpi and 8 wpi).
In Figure 1, we report the number of identified species (R) and the Shannon index. Notably, after the administration of symbiotics, the variability of (R) decreased at 8 wpi across all experimental groups, except the T9M group, where it only decreased in some cases. In contrast, the Shannon index showed greater dispersion following administration; however, no statistical differences were observed between the measurements taken before (4 wpi) and after (8 wpi) the administration of symbiotic.
Quantitatively, comparisons between 4 wpi (before symbiotic administration) and 8 wpi (after symbiotic administration) showed that the R index decreased by approximately 10–15% in T5S and T9S groups, remained stable in T5M, and increased by about 8% in T9M. Similarly, the Shannon index showed a slight increase in the T9M group (0.21 ± 0.04 units), suggesting a mild improvement in microbial evenness following symbiotic supplementation.
Thus, alpha-diversity analysis confirmed similar overall R between groups, with only mild trends toward increased evenness in the moderate injury models.
Although these results were not statistically significant, moderate injuries (particularly T9M) tended to show slightly higher R and Shannon values following symbiotic supplementation, suggesting a mild increase in evenness and species distribution compared with severe injuries.
Relative abundance was calculated, and the most abundant genera per sample are presented below in Figure 2. The most prevalent genera were Bifidobacterium, Lactobacillus, Clostridium, and Alkaliphilus. After treatment (8 wpi), the microbiome composition across groups became more homogeneous. The predominant genera—Bifidobacterium, Lactobacillus, Clostridium, Alkaliphilus, Allobaculum, and Sarcina—together represented approximately 90% of the total abundance.
Quantitative analysis of microbial composition revealed time-dependent changes following symbiotic administration. In the T9M group, Clostridium decreased from 18.6% at 4 wpi to 7.0% at 8 wpi, while Bifidobacterium increased from 32.1% to 34.9%. In parallel, Lactobacillus increased from 17.1% to 32.8%, indicating a shift toward beneficial taxa after treatment.
In the T5M group, Bifidobacterium increased from 30.8% at 4 wpi to 37.1% at 8 wpi, whereas Lactobacillus decreased from 28.7% to 18.5%. Changes in Clostridium were modest, increasing from 9.3% to 12.1%. At the family level, Enterobacteriaceae showed a reduction after symbiotic administration across groups; this taxon is represented accordingly in Figure 2 (see legend).
In severe models (T5S and T9S), compositional shifts were less pronounced, indicating a severity-dependent microbial response. All comparisons correspond to changes between before (4 wpi) and after (8 wpi) symbiotic administration, consistent with the graphical representation.
The Shannon diversity index mirrored these findings, indicating greater evenness in T9M and T5M after symbiotic supplementation.
Before symbiotic administration at 4 wpi, the most abundant genera across groups were Bifidobacterium, Lactobacillus, Clostridium, and Alkaliphilus. After symbiotic administration at 8 wpi, all the groups exhibited a similar microbiome composition where Bifidobacterium, Lactobacillus, Clostridium, Alkaliphilus, Allobaculum, Turicibacter, Natronincola, Sarcina, Akkermansia, Johnsonella, and Blautia together constituted 89.96% of the relative abundance. The eight principal genera of each group were represented in a heat map, as seen in Figure 3.
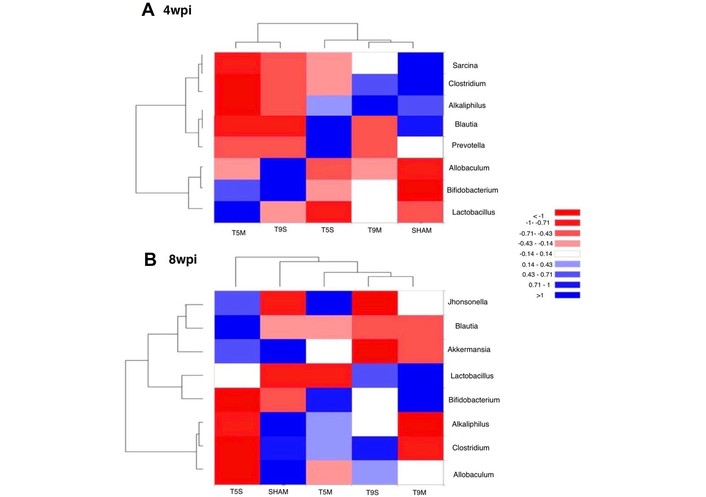
Analysis of species abundance clustering in the heat map. The longitudinal axis shows the group information, while the transverse axis represents the species annotation information. The clustering tree on the left of the figure displays species clustering, and the tree above illustrates the clustering among sample groups. The values in the heat map represent the z-scores obtained after standardization of the relative abundance of species in each row, reflecting gut microbial distribution at the genus level for each experimental group at 4 wpi and 8 wpi. The blue color indicates a higher percentage of relative abundance. wpi: weeks post-injury.
The main genera before and after symbiotic administration were grouped by category, as represented in Table 1, arranged from highest to lowest abundance.
Comparative relative abundance percentages of the gut microbiome before and after symbiotic administration.
| Group | % of the total relative abundance | |
|---|---|---|
| Before symbiotic administration (4wpi) | After symbiotic administration (8 wpi) | |
| Sham | Clostridium, Alkaliphilus, Bifidobacterium, Blautia, Lactobacillus, Sarcina, Turicibacter, and Natronincola (78.46%). | Lactobacillus, Blautia, Bifidobacterium, Alkaliphilus, Clostridium, Ruminococcus, Johnsonella, Akkermansia, Natronincola, and Oscillospira (81.13%). |
| T5S | Bifidobacterium, Clostridium, Alkaliphilus, Prevotella, Blautia, Lactobacillus, Sarcina, and Natronincola (72.87%). | Lactobacillus, Blautia, Bifidobacterium, Alkaliphilus, Clostridium, Ruminococcus, Johnsonella, Akkermansia, Natronincola, and Oscillospira (81.13%). |
| T5M | Bifidobacterium, Lactobacillus, Clostridium, Alkaliphilus, Allobaculum, and Natronincola (83.22%). | Bifidobacterium, Lactobacillus, Clostridium, Alkaliphilus, Allobaculum, Sarcina, and Natronincola (82.18%). |
| T9S | Bifidobacterium, Clostridium, Alkaliphilus, Allobaculum, Lactobacillus, and Blautia (83.56%). | Bifidobacterium, Lactobacillus, Clostridium, Alkaliphilus, Allobaculum, Sarcina, Natronincola, and Turicibacter (91.9%). |
| T9M | Bifidobacterium, Clostridium, Alkaliphilus, Lactobacillus, Allobaculum, Blautia, Sarcina, and Natronincola (84.94%). | Lactobacillus, Bifidobacterium, Clostridium, Alkaliphilus, and Allobaculum (82.99%). |
wpi: weeks post-injury; T5M: thoracic five moderate; T5S: thoracic five severe; T9M: thoracic nine moderate; T9S: thoracic nine severe.
To determine significant dissimilarities in microbiome composition between the experimental groups, PERMANOVA analysis was performed to assess the effects of injury intensity and level. First, we evaluated dissimilarities between the groups before symbiotic administration, which resulted in an explained variation (EV) of 23.5% with a significant difference (p = 0.004). After symbiotic administration, the dissimilarities between the groups showed an EV of 12.7% with a significant difference (p < 0.05), suggesting that symbiotic administration promotes homogenization of microbiota genera irrespective of injury level or severity.
Next, we compared each group to itself before and after symbiotic administration using PERMANOVA, yielding the following results: T5S with an EV of 25.6% (p = 0.2); T5M with an EV of 12.3% (p = 0.439); T9S with an EV of 41.4% (p = 0.058); T9M with an EV of 83.3% (p = 0.035); and Sham with an EV of 45.4% (p = 0.1). The principal observation of EV was noted in the T9 level and Sham. See Table 2.
PERMANOVA analysis with the ten principal genera identifies the different groups.
| Before symbiotic administration | After symbiotic administration | EV and PERMANOVA | ||
|---|---|---|---|---|
| Sham | ||||
| Genus | % | Genus | % | EV and PERMANOVA |
| ClostridiumAlkaliphilusBifidobacteriumSarcinaTuricibacterLactobacillusBlautiaNatronincolaRuminococcusAllobaculum | 27.317.516.17.97.66.25.64.62.31.2 | BifidobacteriumLactobacillusClostridiumAlkaliphilusAllobaculumNatronincolaAkkermansiaTuricibacterSarcinaMesoplasma | 24.216.314.713.79.43.73.53.42.72.0 | 45.4% (p = 0.1) |
| T5S | ||||
| Genus | % | Genus | % | EV and PERMANOVA |
| BifidobacteriumClostridiumAlkaliphilusPrevotellaBlautiaLactobacillusSarcinaNatronincolaBacteroidesAllobaculum | 15.56.85.95.43.83.73.52.11.51.3 | LactobacillusBlautiaBifidobacteriumAlkaliphilusClostridiumRuminococcusJohnsonellaAkkermansiaNatronincolaPelagicoccus | 23.119.515.17.96.94.43.82.52.21.9 | 25.6% (p = 0.2) |
| T5M | ||||
| Genus | % | Genus | % | EV and PERMANOVA |
| BifidobacteriumLactobacillusClostridiumAlkaliphilusAllobaculumBlautiaNatronincolaSarcinaPediococcusParabacteroides | 30.828.79.38.75.12.42.11.81.81.6 | BifidobacteriumLactobacillusClostridiumAlkaliphilusAllobaculumSarcinaNatronincolaAkkermansiaPediococcusTuricibacter | 37.118.512.19.75.22.82.41.71.41.4 | 12.3% (p = 0.439) |
| T9S | ||||
| Genus | % | Genus | % | EV and PERMANOVA |
| BifidobacteriumClostridiumLactobacillusAllobaculumAlkaliphilusBlautiaSarcinaNatronincolaMesoplasmaOscillospira | 41.413.310.29.99.42.92.51.81.31.1 | BifidobacteriumLactobacillusClostridiumAlkaliphilusSarcinaAllobaculumNatronincolaTuricibacterPediococcusBlautia | 26.521.919.710.47.04.72.82.21.20.9 | 41.4% (p = 0.058) |
| T9M | ||||
| Genus | % | Genus | % | EV and PERMANOVA |
| BifidobacteriumClostridiumLactobacillusAlkaliphilusAllobaculumSarcinaNatronincolaRuminococcusPediococcusTuricibacter | 32.118.617.116.83.92.92.31.11.00.7 | BifidobacteriumLactobacillusClostridiumAlkaliphilusAllobaculumPediococcusBlautiaSarcinaRuminococcusMesoplasma | 34.932.87.06.74.71.71.51.51.31.0 | 83.3% (p = 0.035) |
EV: explained variation; T5M: thoracic five moderate; T5S: thoracic five severe; T9M: thoracic nine moderate; T9S: thoracic nine severe.
Before symbiotic administration (4 wpi), butyrate levels varied across experimental groups and did not consistently decrease in all SCI conditions compared with the Sham group. While some injury groups showed lower concentrations, others (e.g., T5S) exhibited comparable or even higher baseline levels. In contrast, animals with moderate T9 injury showed relatively higher baseline concentrations overall (Figure 4).
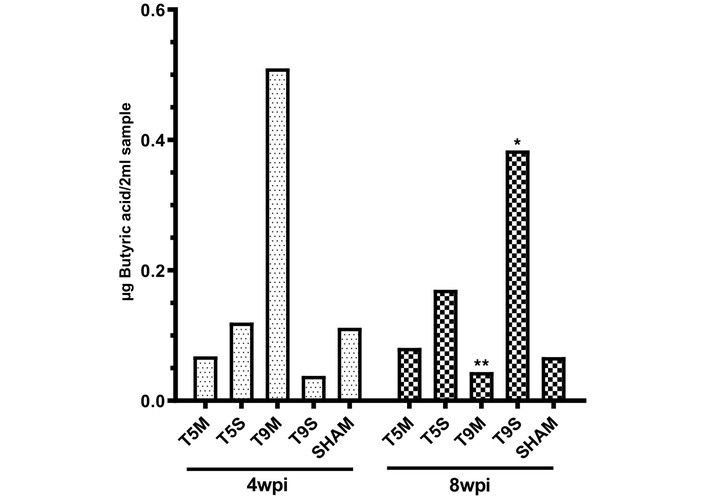
Butyric acid concentration before and after symbiotic administration. Bar graph showing butyric acid levels in pooled fecal samples from each experimental group at 4 wpi (before symbiotic administration) and 8 wpi (after symbiotic administration). Values are expressed as mean ± SD of technical replicates. Statistical significance is indicated as follows: * p < 0.05, ** p < 0.01. Comparisons correspond to within-group differences between 4 wpi and 8 wpi. wpi: weeks post-injury; SD: standard deviation.
After four weeks of symbiotic supplementation (8 wpi), butyrate concentrations showed a heterogeneous response across experimental groups. While some groups (e.g., T9S) exhibited an increase, others, such as T9M, showed a decrease. Changes in T5 groups were modest, whereas the Sham group remained relatively stable across both time points. A significant increase in butyrate concentration was observed in the T9S group (p < 0.05), whereas other groups showed no consistent significant changes.
Overall, these results suggest that symbiotic supplementation may enhance butyrate production, particularly in animals with moderate T9 injury.
To verify the consistency of the contusion model, we evaluated motor recovery in all groups following injury. A mixed-effects model (REML) with Tukey’s multiple comparisons test was used to assess motor recovery over time, comparing the period before (4 wpi) and after (8 wpi) symbiotic administration.
During the first four weeks, we observed the expected spontaneous recovery of locomotor function across all groups. However, following treatment, this improvement continued to progress, despite the lesion having reached its chronic phase.
The recovery was statistically significant in the T9M (p < 0.0001), T9S (p = 0.0166), and T5M (p = 0.0013) groups (Figure 5).
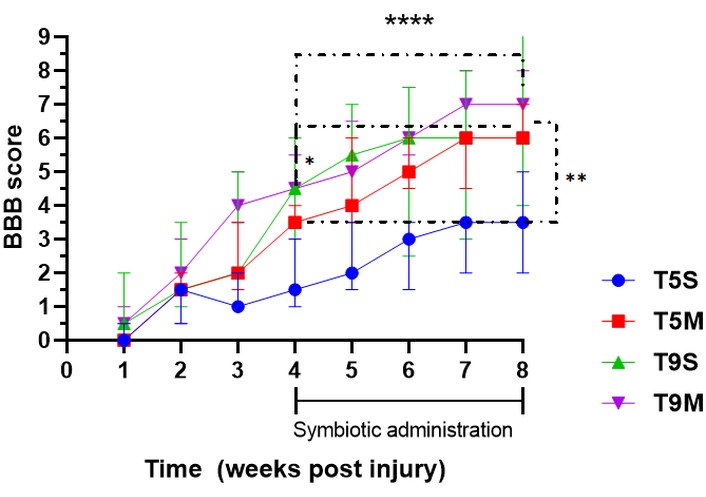
Motor recovery throughout the experiment. Statistical analysis was evaluated with REML with Tukey’s multiple comparisons to evaluate all animals per group over time. Values are expressed as mean ± SD. Significant improvements were observed during the symbiotic administration period, particularly in T9 groups. Asterisks indicate statistically significant differences as follows: * p < 0.05, ** p < 0.01, **** p < 0.0001. Specifically, * indicates within-group differences during the early phase of treatment (around week 4–5), ** indicates significant differences between groups at week 8 (notably between T5S and treated groups), and **** indicates highly significant improvements across time points during the symbiotic administration period (4–8 wpi), particularly in T9M and T9S groups. wpi: weeks post-injury; REML: mixed-effects model; SD: standard deviation; T5S: thoracic five severe; T9M: thoracic nine moderate; T9S: thoracic nine severe.
Overall, the symbiotic formulation with E. faecium and agave inulin promoted compositional and metabolic restoration of the gut microbiota primarily in moderate injuries. However, since no untreated SCI controls were maintained at 8 weeks, the observed effects should be interpreted cautiously as potential improvements rather than definitive treatment effects.
Although the study of the microbiota-intestine-brain axis continues to shed light on the symbiotic dynamics that allow us to maintain homeostasis with our external environment, our current knowledge remains limited. This project aimed to identify how the level and intensity of SCI influence the composition of the intestinal microbiota and butyrate levels in feces. Injuries at the T5 level, although less commonly studied in SCI models, are known to cause more severe physiological consequences than injuries at T9. This study evaluated two scenarios: a) a T5 lesion, which interrupts extrinsic innervation of the small and large intestine, rectum, and spleen without affecting ganglia from other plexuses such as the cardiac plexus, and b) a T9 lesion, affecting only the extrinsic innervation to the colon and rectum.
SCI has been shown to negatively affect the gut microbiota, as described by other authors [17, 27]. In our study, microbial species diversity was most affected in the T5S group at four wpi. By eight weeks, intergroup variation decreased, while intragroup variation increased, although the total number of identified species and variability showed an overall decreasing trend.
Notably, a general increase in Bifidobacterium was observed across all injured groups four weeks post-SCI. In contrast, Clostridium accounted for 27.28% of the microbiota in the Sham group, but remained below 20% in all other experimental groups. The T5S group displayed the most pronounced alterations, with many genera appearing in less than 50% of the samples, indicating significant disruption in microbial stability. The impact of SCI level and severity on microbial recovery was further evidenced after symbiotic administration. Two main trends were observed. 1. In all groups except Sham, there was a homogenization in the relative abundance of bacterial species, leading to greater similarity between groups. 2. This effect was absent in the T5S group, which maintained a distinct profile. In most treated groups, Bifidobacterium, Clostridium, and Lactobacillus predominated, suggesting a partial return to eubiosis [15]. However, in T5S, the dominant genera were Blautia and Lactobacillus, indicating a persistent imbalance likely due to greater neural disruption. These findings align only partially with previous studies. For instance, Lin et al. [27] (2020) reported significant alterations in the fecal microbiota composition of patients with SCI, including changes in the relative abundance of several bacterial taxa associated with gut dysbiosis. It is important to note that interpretation of microbial changes at the genus level should be made with caution, as some genera, such as Clostridium and Bifidobacterium, contain both beneficial and pathogenic species. The increase in Clostridium observed after symbiotic treatment in our study likely reflects the enrichment of beneficial butyrate-producing species (e.g., C. butyricum), consistent with the concomitant rise in fecal butyrate levels. Similarly, the elevation in Bifidobacterium abundance may correspond to probiotic strains that contribute to carbohydrate fermentation and epithelial health. The differences with previous reports may relate to variations in injury level, severity, post-injury phase, or sequencing resolution. Our analysis was based on 16S rRNA V3–V4 region sequencing, which provides genus-level but not species-level identification; therefore, the results should be interpreted as indicative of functional recovery rather than exact taxonomic reconstitution.
Hierarchical clustering analyses supported some of these differences in microbial composition, although not all findings were statistically significant. It is worth noting that such predictive models rely on dissimilarity measures and do not identify individual taxa responsible for group differences [22, 23]. In clinical studies, dysbiosis has been reported to be more severe in upper motor neuron injuries. Predominant genera in lesion groups include Blautia, Bifidobacterium, Faecalibacterium, and Ruminococcus, while Roseburia, Pseudobutyrivibrio, Dialister, and Megamonas are often depleted [15, 27]. These patterns are consistent with findings in the T5-injured animals of our study.
Regarding butyrate levels, the data suggest that T5-level injuries may impair the assimilation of symbiotics, likely due to disruptions in gut innervation. Although fecal pH was not directly measured in this study, it is known that butyrate-producing bacteria contribute to the acidification of the colon through SCFA production. Based on this, one could hypothesize that animals in the Sham, T9S, and T9M groups might present a more acidic fecal environment, whereas those in the T5S and T5M groups might exhibit a more alkaline profile, consistent with a lower abundance of butyrogenic bacteria. These suppositions, however, remain speculative and highlight the need for future studies to include direct fecal pH measurements, which could serve as a functional readout of SCFA activity and microbiota composition in SCI models. Assessing pH alongside microbiota profiling and SCFA quantification (e.g., butyrate, acetate, propionate) would provide a more comprehensive understanding of gut environment alterations following SCI.
Notwithstanding, one limitation of our study was the pooling of fecal samples, which may have masked individual variability. Ideally, fecal microbiota analysis should be performed per subject to improve resolution and accuracy [28]. Additionally, the relatively small sample size (n = 4 per group) may limit the generalizability of the findings and reduce the ability to detect subtle inter-individual differences. Despite these constraints, the observed microbial and metabolic trends were consistent across experimental groups, supporting the robustness of our findings. Another limitation of this study is the absence of additional untreated SCI cohorts maintained until 8 wpi. However, the longitudinal pre- or post-test design allowed each animal to serve as its own control, reducing biological variability. The Sham group provided the physiological reference for baseline microbial and metabolic conditions. Moreover, the study design did not include SCI groups that remained untreated at eight wpi. As a result, it is difficult to fully distinguish the effects of the symbiotic treatment from the natural progression of recovery or time-dependent microbial fluctuations. This limitation affects the interpretation of metagenomic, butyrate, and locomotor outcomes, which should therefore be regarded as indicative of potential functional improvement rather than definitive proof of treatment efficacy. Future studies should incorporate untreated SCI controls at equivalent time points to better isolate the specific contribution of the symbiotic intervention.
Looking more closely at Clostridium, a well-established butyrate producer, its abundance was reduced in all experimental groups at four weeks. After symbiotic therapy, Bifidobacterium, Lactobacillus, and Clostridium populations increased, yet the T5S group again showed limited improvement, potentially due to greater impairment of the gut-brain axis [29–31].
To better understand the distinct roles of SCFAs, future studies should include quantification of other SCFAs, such as acetate and propionate, to differentiate their contributions to gut and systemic homeostasis. From a functional perspective, the motor recovery patterns observed via BBB scoring reflected what is well-established in SCI models. Rats with lower-level injuries (T9) exhibited better locomotor recovery than those with higher-level injuries (T5). Similarly, animals with moderate injuries recovered more than those with severe injuries, confirming the well-documented correlation between injury severity, lesion level, and functional outcome. These differences became especially clear by week 8, with the T9M group showing the greatest improvement, followed by T5M, T9S, and finally T5S. These findings are consistent with the natural progression of SCI and emphasize that lesion level and intensity are key determinants of motor function recovery. Furthermore, discrepancies among studies reporting divergent trends in Bifidobacterium and Clostridium abundance following SCI (e.g., Jeffrey et al. [17], 2018 vs. Kigerl et al. [28], 2016) may be explained by methodological heterogeneity, including differences in post-injury sampling windows, dietary composition, and the use of antibiotics or fiber sources, as highlighted by Gungor et al. [15] (2016). Integrating these contextual factors provides a more coherent framework for understanding microbial variability across experimental models.
In summary, this study provides important insights into how the level and severity of SCI influence gut microbiota composition and butyrate metabolism. The microbial alterations were most profound in animals with high-level and severe injuries (T5S), which also exhibited less favorable bacterial recovery. Meanwhile, motor function improved in line with expectations, with better outcomes in lower and moderate injuries, independent of symbiotic intervention.
These findings underscore the need to consider injury-specific factors—especially neural disruption and gastrointestinal innervation—when interpreting microbial and physiological outcomes in SCI. Future research should focus on individual-level microbiota analyses, early interventions, and mechanistic studies exploring SCFA-specific effects on host health.
The present study demonstrates that symbiotic administration with E. faecium and agave inulin significantly influenced gut microbiota composition and metabolic function following SCI. Treatment led to the recovery of beneficial bacterial genera such as Bifidobacterium, Clostridium, and Lactobacillus. Locomotor recovery followed the expected pattern based on lesion level and injury severity, with better outcomes observed in moderate and lower thoracic injuries. Separately, the microbial and metabolic changes observed after symbiotic administration suggest a potential modulatory effect on the microbiota-gut-spinal cord axis. However, the observed changes in butyrate concentration were inconsistent with the expected relationship between butyrate levels and injury severity.
Although limited by the small sample size and pooled sequencing design, the consistent post-treatment trends indicate that symbiotic administration exerted a restorative and modulatory effect on intestinal microbial ecology and SCFA metabolism. These findings underscore the therapeutic potential of symbiotic supplementation as a complementary strategy to improve gastrointestinal and neurological function after SCI. Future studies with larger cohorts and individual-level analyses will help confirm these effects and explore their underlying mechanisms.
16S rRNA: 16S ribosomal RNA
AEH: Alimentos Esenciales para la Humanidad
BBB: Basso, Beattie, and Bresnahan
BD: bowel dysfunction
EV: explained variation
GC: gas chromatography
GIT: gastrointestinal tract
OTUs: Operational Taxonomic Units
PCoA: principal coordinate analysis
R: Richness
REML: mixed-effects model
SCFAs: short-chain fatty acids
SCI: spinal cord injury
SD: standard deviation
T5M: thoracic five moderate
T5S: thoracic five severe
T9M: thoracic nine moderate
T9S: thoracic nine severe
TGF-β: transforming growth factor-beta
wpi: weeks post-injury
EGV: Conceptualization, Investigation, Writing—review & editing, Visualization, Formal analysis. EDlCC: Investigation, Writing—original draft. RRB: Investigation. AFR: Investigation. TPL: Validation. DRMQ: Investigation, Validation. HFNV: Supervision. IIM: Investigation, Writing—review & editing. MAVH: Supervision. EMAM: Supervision. APIG: Writing—review & editing. AI: Conceptualization, Writing—review & editing, Validation, Supervision. All authors read and approved the submitted version.
Antonio Ibarra, who is the Editorial Board Member and Guest Editor of Exploration of Neuroprotective Therapy, had no involvement in the decision-making or the review process of this manuscript. The other authors declare no conflicts of interest.
All experimental procedures were conducted in accordance with protocols approved by the National Institutes of Health (US) guidelines and the Guide for the Care and Use of Animals. The Animal Bioethics and Welfare Committee of the Universidad Anáhuac México approved all animal procedures (Registry number: 201702-CI) in accordance with the National Institutes of Health Guide for the care and use of laboratory animals, and the Mexican Official Norm on Principles of Laboratory Animal Care (NOM 062-ZOO-1999).
Not applicable.
Not applicable.
The data supporting the findings will be made available upon request.
This study was supported by Universidad Anáhuac México, Grant/Award Number: 201702; Universidad Anáhuac Mexico, Grant/Award Number: 201707. The funder(s) had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 426
Download: 14
Times Cited: 0
Adnan Akhtar Shaikh ... Niveditha Nair
Salomón Páez-García ... Miguel Germán Borda
Zhengrui Li ... Jing Li
Diego Fernández-Lázaro ... Juan Mielgo-Ayuso
Ericka C. Loza López ... Felipe Esparza Salazar
Fernando Leal-Martínez, Irene Alejandra Berumen Coronado
Alejandro Borrego-Ruiz, Juan J. Borrego
Natalia Rojas-Sánchez ... Antonio Ibarra
