Review
Review

Affiliation:
1School of Pharmaceutical Sciences, Faculty of Pharmacy, IFTM University, Moradabad 244102, Uttar Pradesh, India
Email: AKarvindsr01@gmail.com
Affiliation:
1School of Pharmaceutical Sciences, Faculty of Pharmacy, IFTM University, Moradabad 244102, Uttar Pradesh, India
Affiliation:
2Raj Kumar Goel Institute of Technology (Pharmacy), Ghaziabad 201003, Uttar Pradesh, India
Affiliation:
3SRM Modinagar College of Pharmacy, Faculty of Medicine & Health Sciences, SRM Institute of Science and Technology, NCR Campus, Delhi-NCR Campus, Ghaziabad 2012024, Uttar Pradesh, India
Affiliation:
4Department of Pharmaceutics, Moradabad Educational Trust, Group of Institutions, Faculty of Pharmacy, Moradabad 244001, Uttar Pradesh, India
Affiliation:
5Department of Pharmaceutical Chemistry, Moradabad Educational Trust, Group of Institutions, Faculty of Pharmacy, Moradabad 244001, Uttar Pradesh, India
Affiliation:
6SOS School of Pharmacy, Faculty of Pharmacy, IFTM University, Moradabad 244102, Uttar Pradesh, India
Affiliation:
7Department of Biosciences, Saveetha School of Engineering, Saveetha Institute of Medical and Technical Sciences, Chennai 602105, Tamil Nadu, India
Affiliation:
8Centre for Research Impact & Outcome, Chitkara College of Pharmacy, Chitkara University, Rajpura 140401, Punjab, India
Email: chopraontheride@gmail.com
ORCID: https://orcid.org/0000-0001-8867-7603
Explor Foods Foodomics. 2026;4:1010174 DOI: https://doi.org/10.37349/eff.2026.1010174
Received: March 04, 2026 Accepted: May 15, 2026 Published: July 02, 2026
Academic Editor: Cristina Garcia Viguera, CEBAS-CSIC, Spain
Apigenin, a dietary flavonoid that occurs naturally in parsley, chamomile, and a variety of other plant foods, has attracted increasing scientific interest for its broad spectrum of pharmacological effects, such as antioxidant, anti-inflammatory, anticancer, antimicrobial, neuroprotective, and cardioprotective activities. Being structurally related to quercetin, apigenin exhibits significant therapeutic potential; however, its clinical application is limited by poor aqueous solubility and low bioavailability. Recent studies have investigated the synergistic ability of apigenin when it is used in combination with a variety of small-molecule agents to overcome these challenges and improve therapeutic efficacy. Such combinations have been shown to be effective in the management of cancer, neurodegenerative disorders, and metabolic syndromes through mechanisms that include modulation of oxidative stress, cell cycle arrest, induction of apoptosis, and interference with major signaling pathways like PI3K/Akt, NF-κB, and MAPK. This review uniquely focuses on drug-specific synergistic interactions between apigenin and conventional small-molecule therapeutics, highlighting mechanistic pathways such as PI3K/Akt, NF-κB, MAPK, and drug transporter modulation. By critically analyzing these interactions, the study provides insights into combination-based therapeutic strategies and identifies key gaps for clinical translation. The inclusion criteria comprised studies published between 2000 and 2025, written in English, focusing on the pharmacological activity of apigenin. Electronic academic databases like PubMed, IEEE Xplore, Scopus, and ScienceDirect that provide extensive access to peer-reviewed medical and technological studies were the primary source of literature reviewed in this study. Keywords like “pharmacological evaluation,” “synergistic effects,” and “apigenin” were used to choose articles. This search strategy enables the identification of relevant original studies and review articles addressing the therapeutic potential of apigenin.
In recent years, there has been significant interest in natural extracts and compounds that offer health benefits, with polyphenols as a prominent example. Currently, over 8,000 polyphenol compounds have been identified, with flavones being the most abundant among them. Flavones have garnered considerable attention due to their diverse biochemical and pharmacological effects. Originating as the products of secondary metabolism in plants, flavones are widely present in various plant foods that have been extensively studied for more than a century. The biological activity of flavones was first explored by Rusnak and Szent-Gyorgyi in 1936 [1]. Since it is a naturally occurring flavone, apigenin can be found in abundance in many plant foods. It is found abundantly in foods like herbs (parsley, chamomile, oregano, and basil), fruits and vegetables (celery, onions, oranges, and tea). Since apigenin is found in large amounts in foods we consume daily from plants, it is one of the significant flavonoids in our diet [2]. Chamomile tea is one of the richest dietary sources and is a common and readily available form of apigenin consumption. Because it is found in such common foods, apigenin is a major contributor to the health benefits of a diet rich in plants. The major dietary sources of apigenin are illustrated in Figure 1 in the following article.

Major natural dietary sources of apigenin include parsley, chamomile, oregano, basil, celery, onions, and oranges.
The chemical name of apigenin is 4′,5,7-trihydroxyflavone. However, its poor aqueous solubility significantly reduces oral bioavailability, thereby limiting its pharmaceutical applications and restricting its usage in the food and pharmaceutical industry [3]. Flavonoids are a diverse class of polyphenolic compounds found abundantly in plant tissues, where they play crucial roles in defense, metabolism, and reproduction. With over 6,000 identified compounds, flavonoids exhibit structural complexity based on their flavone nucleus and substitution patterns, leading to various subclasses such as flavones, flavonols, flavanones, flavanonols, and anthocyanidins, as shown in Figure 2. Their multifunctional nature extends beyond plant physiology, as they contribute to human health through their antioxidant, anti-inflammatory, and potential stress-relieving properties. Given their biological significance, flavonoids continue to be a subject of extensive research in pharmacology, nutrition, and functional food development [4].
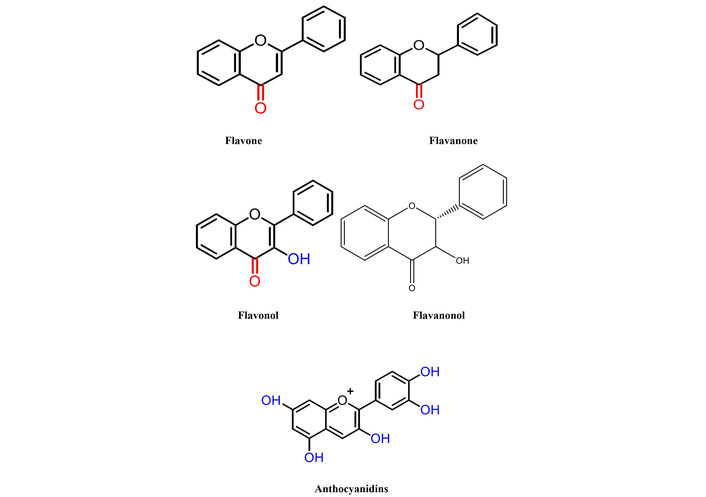
Structural classification of flavonoids. The flavone subclass, which includes apigenin, is distinguished by a double bond between C2–C3 and a ketone group at C4.
Flavones, which occur mainly in plants such as 7-O-glycosides and C-glycosides, exhibit structural variability due to changes in glycosylation and other attached functional groups. The structural variations of flavones, including apigenin, are illustrated in Figure 3. O-glycosides, on the other hand, are hydrolysable before analysis, whereas C-glycosides are stable and need special analytical techniques. Both glycosides and aglycones are considered when quantifying flavones in food studies, and daily consumption is reported variably across populations. These differences underline the potential benefits of flavones for human nutrition and health and stress the significance of using consistent approaches in flavone research [5].
This review endeavors to present an integrated perspective on the multiplicity of pharmacological activities of apigenin, encompassing its cardioprotective, neuroprotective, anti-inflammatory, and anticancer activities. Being a flavonoid of natural origin, apigenin possesses great therapeutic potential across numerous therapeutic categories; nevertheless, its clinical application is severely limited by its poor bioavailability. The current paper also focuses on the restriction of supplementation of apigenin and its rarity in typical food sources. Further, it highlights the therapeutic potential of the compound to elicit enhanced efficacy by synergistic interactions with other small-molecule drugs. The review also considers apigenin’s safety based on preclinical and clinical data to consider any associated side effects. Lastly, it identifies major knowledge gaps in current research and offers recommendations for future studies, particularly in areas such as new drug delivery systems, improved bioavailability, and clinical efficacy of apigenin therapy. This review first provides a concise overview of the pharmacological profile of apigenin to establish a mechanistic foundation, which is subsequently used to contextualize its synergistic interactions with conventional small-molecule drugs.
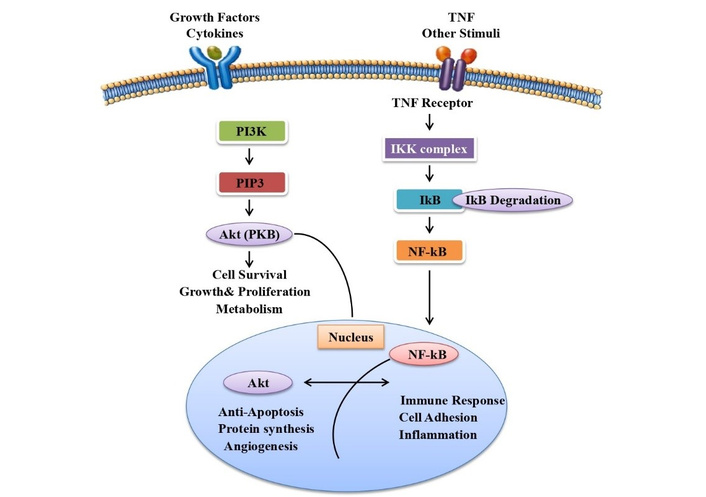
To ensure consistency with the primary objective of this review, only studies with direct relevance to apigenin were included, particularly those addressing its chemical structure, pharmacological activities, and underlying molecular mechanisms. Eligible studies comprised original research articles employing in vitro, in vivo, clinical, or in silico approaches that specifically investigated the biological effects of apigenin, such as antioxidant, anti-inflammatory, anticancer, neuroprotective, and cardioprotective activities. Special emphasis was placed on studies elucidating mechanistic pathways, including but not limited to PI3K/Akt, NF-κB, MAPK, and apoptosis-related signaling. Additionally, studies exploring synergistic, additive, or potentiating interactions between apigenin and small-molecule therapeutic agents were included to support the central theme of combination-based therapy.
Studies involving other flavonoids were considered only when they provided direct comparative insights or mechanistic relevance to apigenin, particularly in terms of structural similarity or shared biological pathways. Only peer-reviewed articles published in English between 2000 and 2025 were included, with preference given to full-text studies available through established academic databases such as PubMed, Scopus, ScienceDirect, and IEEE Xplore. All selected studies were critically evaluated for methodological quality, direct relevance to apigenin, and traceability to original experimental or clinical findings. A schematic representation (Figure 4) summarizes the molecular mechanisms and pharmacokinetic behavior of apigenin, providing a foundational understanding of its bioavailability and therapeutic interactions, which supports the inclusion criteria applied in this review.
Studies were excluded if they did not align with the central focus of this review on apigenin’s structure, biological activities, and mechanisms of action. Specifically, studies primarily investigating flavonoids other than apigenin were excluded unless they offered clear comparative or mechanistic relevance to apigenin. Research involving crude plant extracts without proper isolation, identification, or quantification of apigenin was also excluded to maintain specificity. Studies lacking mechanistic insights or those not directly addressing the pharmacological effects of apigenin were considered ineligible. Furthermore, studies evaluating apigenin without examining its biological activity or its interaction with other therapeutic agents were excluded unless they contributed substantial mechanistic understanding. Research focused solely on combinations with macromolecules such as proteins, peptides, antibodies, or nanoparticles was excluded unless small-molecule interactions were clearly demonstrated. Non-research articles, including editorials, commentaries, conference abstracts, and reports with insufficient experimental data, were also excluded, along with publications in languages other than English.
Overall, current evidence supporting the therapeutic application of apigenin is predominantly derived from in vitro and animal studies, with limited clinical validation. Additional challenges include poor aqueous solubility and bioavailability, variability in commercial formulations, insufficient standardization of dosing strategies, and inadequate understanding of long-term safety and drug–drug interactions. Furthermore, gender-specific responses and interindividual genetic variability remain poorly characterized. Addressing these limitations through well-designed clinical trials, standardized formulations, pharmacokinetic investigations, and mechanistic studies will be essential for translating the promising preclinical findings into clinically applicable therapeutic strategies.
Apigenin, a naturally occurring flavonoid, and its derivatives exhibit a wide range of pharmacological activities. These bioactivities contribute to its potential use in disease prevention and treatment. These antioxidant properties may contribute to enhanced efficacy when apigenin is combined with chemotherapeutic agents by reducing oxidative stress-mediated drug resistance.
Pancreatic cancer (PC) has a five-year survival rate of fewer than 5%, making it the fourth most common cause of cancer-related death [6]. Although there have been improvements in therapeutic approaches, mortality is still not significantly improved by the aggressive nature of PC and its resistance to existing therapy. Conventional modalities of therapy, including chemotherapy, surgery, and radiotherapy, have mostly limited success, and resistance in cancer cells is common. To counter this issue, combination chemotherapy with anti-tumor drugs has been investigated [7]. Natural plant-derived compounds have gained attention for their ability to enhance chemotherapy sensitivity, suppress tumor progression, and target multiple molecular pathways involved in cancer resistance. Apigenin, a naturally occurring flavonoid, has shown promising anti-tumor effects in various cancers, including PC [8]. It regulates major oncogenic signaling pathways such as Akt, Wnt, and Nrf2 and can inhibit epithelial-to-mesenchymal transition (EMT) and thereby inhibit metastasis and cancer cell proliferation. From studies, apigenin, similar to other phytochemicals, curcumin, has the potential to enhance the effectiveness of chemotherapy and inhibit tumor aggressiveness by inhibiting processes that benefit cancer cells. Due to its multiple-targeting potential and low toxicity, apigenin is the ideal natural compound for use in combination therapy against PC. Further studies need to be done to enhance its bioavailability and therapeutic applications [9, 10]. Apigenin suppressed adenoid cystic carcinoma cell growth by downregulating glucose transporter-1 (GLUT-1) expression, implying a metabolic interference anticancer process. In BCPAP cells, apigenin caused autophagic cell death, implying that apigenin kills thyroid cancer cells by modulating autophagy. Apigenin inhibited the proliferation and invasion of osteosarcoma cells by suppressing the Wnt/β-catenin signaling pathway, a critical pathway in bone cancer progression. Apigenin enhanced the therapeutic efficacy of TRAIL (TNF-related apoptosis-inducing ligand) in non-small cell lung cancer by upregulating death receptors DR4 and DR5 in a p53-dependent manner, suggesting a synergistic approach in lung cancer therapy [11]. In MDA-MB-231 breast cancer cells, apigenin induced G2/M phase arrest and promoted histone H3 acetylation-mediated p21 (WAF1/CIP1) expression, leading to inhibited cell proliferation and tumor growth. Apigenin inhibited the proliferation of renal cell carcinoma cells by causing DNA damage and inducing G2/M phase cell cycle arrest through modulation of the ATM signaling pathway [12]. Apigenin exhibited cytotoxic effects on malignant mesothelioma cells in a dose- and time-dependent manner. Treatment led to increased reactive oxygen species (ROS) generation, DNA damage, upregulation of p53, and activation of apoptotic pathways, suggesting its potential in mesothelioma therapy [13]. These mechanistic effects provide a strong basis for synergistic interactions with chemotherapeutic agents such as doxorubicin and cisplatin, where apigenin enhances drug accumulation, modulates apoptosis pathways, and overcomes resistance mechanisms.
Apigenin isolated from Patrinia villosa exhibited anti-proliferative effects towards HCC cell growth. Apigenin induced G1 arrest of HepG2 cells, a cell line of liver cancer, possibly by activating the p38 MAPK-p21 signaling pathway. The process also involved control of the CyclinD1-CDK4 complex, the main figure involved in cell cycle progression. These findings suggest that apigenin is an extremely effective therapeutic drug against HCC through control of cell cycle progression and cancer cell growth [14]. Apigenin treatment of glioblastoma cells (U87MG and T98G) induced the generation of ROS, activation of the p38 MAPK and JNK pathways, intracellular calcium elevation, and apoptosis. Notably, apigenin did not kill normal astrocytes, reflecting its glioblastoma cell-selective cytotoxicity. The results suggest that apigenin may serve as an effective glioblastoma drug with decreased effect on normal cells. These findings strongly support the role of apigenin as a chemosensitizing agent in cancer therapy. Its ability to modulate apoptosis, oxidative stress, and drug resistance pathways highlights its potential for synergistic use with standard chemotherapeutic agents such as doxorubicin and cisplatin, thereby improving therapeutic outcomes and reducing resistance [15].
Research isolated apigenin from Portulaca oleracea L. and tested its antibacterial activity against some pathogenic bacteria. The findings showed strong inhibition zones for Salmonella typhimurium and Proteus mirabilis, indicating strong antibacterial activity. This research did not specifically test apigenin’s activity towards Mycobacterium tuberculosis or other mycobacterial species.[16]. The antimycobacterial properties of flavonoids indicated that some flavonoids, like taxifolin, had strong inhibitory action against Mycobacterium tuberculosis. On the other hand, apigenin was found to possess merely moderate to weak mycobacterial activity, indicating that although it does bear some antimycobacterial activity, it is perhaps not as powerful as other flavonoids in this aspect [17]. Also, studies on Schinus terebinthifolius considered the effects of its methanol extract, a particular fraction (A3), and pure apigenin on Mycobacterium bovis BCG growth. The investigation concluded that although the extract and fraction A3 showed some inhibitory activity, pure apigenin showed reduced effect, thus suggesting that other components present in the extract could be accountable for observed antimycobacterial activity. Although apigenin alone exhibits limited antimycobacterial activity, its moderate efficacy suggests potential for synergistic application. Future studies should explore its use in combination with first-line anti-tubercular drugs to enhance efficacy and overcome resistance mechanisms [18].
An examination was made on the impact of apigenin on isolated rat hearts with ischemia/reperfusion (I/R) injury. The findings showed that treatment with apigenin enhanced the cardiac functional recovery, reduced the infarct size in the myocardium, lowered activities of creatine kinase isoenzyme and lactate dehydrogenase in coronary flow, and lowered apoptotic cardiomyocytes. These were linked with the inhibition of the p38 MAPK signaling pathway [19]. Research assessed the effects of apigenin on renovascular hypertension-evoked cardiac hypertrophy in rats. Administering apigenin led to lowered blood pressure, heart weight, cardiomyocyte cross-sectional area, serum angiotensin II, and myocardial free fatty acids. The study indicated that the effects were mediated via down-regulating hypoxia-inducible factor (HIF)-1α and regulating myocardial glucolipid metabolism [20]. Another study investigated apigenin’s cardioprotective activities on diabetic rats with isoproterenol-induced myocardial infarction. Apigenin therapy enhanced hemodynamic parameters, lowered oxidative stress, and lessened apoptosis in myocardial tissue. These responses were attributed to the activation of the peroxisome proliferator-activated receptor-gamma (PPAR-γ) pathway [21]. Apigenin’s effect on mitigating endotoxin-induced myocardial injury in mice was studied. The results showed that apigenin inhibited the expression of pro-inflammatory cytokines, decreased markers of oxidative stress, and blocked NF-κB nuclear translocation. Apigenin also regulated autophagy-related proteins, implying an overarching protective mechanism for cardiac injury induced by endotoxins [22]. A systematic review highlighted apigenin’s protective roles against cardiometabolic diseases. The review emphasized apigenin's ability to reduce oxidative stress, inflammation, and apoptosis, and to improve glucose and lipid metabolism [23]. The researchers proposed that although preclinical results are promising, clinical trials are necessary to establish the human dosage and bioavailability of apigenin. Recent studies discussed the benefits of apigenin in the prevention of cardiac hypertrophy and heart failure. The research explained that apigenin inhibits cardiac hypertrophy through NADPH oxidase-dependent ROS suppression and HIF-1α down-regulation, enhancing myocardial metabolism and function. These cardioprotective effects suggest that apigenin may serve as an adjunctive agent in cardiovascular therapy. Its antioxidant and anti-inflammatory properties could synergistically enhance the efficacy of standard cardioprotective drugs while reducing oxidative stress-mediated damage [24].
An experiment examined apigenin’s action on UVA-induced skin aging. The results confirmed that apigenin recovered the viability of human dermal fibroblasts that had been treated with UVA, lowered cellular senescence, and diminished the expression of matrix metalloproteinase-1 (MMP-1), which breaks down collagen. Clinical use of cream with an apigenin component proved to enhance the density of dermis, improve the elasticity of skin, and lessen fine wrinkles, promising apigenin as an anti-aging cosmetic product [25]. Study on the anti-aging protective effects of apigenin in a D-galactose-induced mouse model of aging. Supplementation with apigenin greatly enhanced behavioral deficits, decreased histopathological injury, and lowered cellular senescence and oxidative stress markers. The study concluded that the anti-aging effects of apigenin could be exerted by induction of the Nrf2 pathway, leading to increased expression of antioxidant enzymes [26]. Studies investigated the protective role of apigenin in a D-galactose-induced model of aging in mice. Supplementation with apigenin markedly ameliorated behavioral impairments, lessened histopathological damage, and minimized markers of cellular senescence and oxidative stress. The authors concluded that apigenin’s anti-aging impact could be mediated by the activation of the Nrf2 pathway, which promotes antioxidant enzyme expression. The anti-aging effects of apigenin, particularly through Nrf2 pathway activation, indicate its potential for synergistic use with other antioxidants or dermatological agents. Such combinations may enhance skin protection and delay aging-related cellular damage more effectively than monotherapy [27].
Apigenin’s effects on cognitive performance in APP/PS1 transgenic mice, an Alzheimer’s disease (AD) model, were studied. The findings showed that apigenin enhanced learning and memory impairment, inhibited amyloid-beta (Aβ) accumulation by suppressing BACE1 and β-CTF expression, improved oxidative stress, and restored the ERK/CREB/BDNF neurotrophic pathway in the cerebral cortex. These observations indicate apigenin’s potential to improve cognitive impairment associated with AD [28]. A study proved that apigenin preserved neurovascular coupling in mice from amyloid-β25-35 toxicity, a peptide linked to AD pathology [29]. A transgenic Drosophila model of AD was used in a study that discovered apigenin treatment caused a dose-dependent reduction in oxidative stress, delayed climb loss, inhibition of acetylcholinesterase activity, and blocking of amyloid-β42 aggregation [30]. Another investigation reported that apigenin restored memory function in mice subjected to chronic unpredictable mild stress. The flavonoid improved cognitive performance reduced oxidative stress markers, normalized serum corticosterone levels, and upregulated BDNF, pERK, and pCREB expressions in the prefrontal cortex and hippocampus [31]. Researchers examined apigenin’s neuroprotective effects in neonatal rats subjected to hypoxic-ischemic brain injury. The study found that apigenin significantly reduced infarct volume, decreased cerebral edema, suppressed inflammatory responses, inhibited apoptosis, and promoted tissue structure recovery. Mechanistically, these benefits were attributed to the activation of the PI3K/Akt/Nrf2 signaling pathway, highlighting apigenin’s therapeutic potential for neonatal brain injuries [32]. The preclinical investigations analyzed apigenin’s effects on cognitive and neurobehavioral dysfunctions. The review summed up that apigenin augments learning and memory, enhances locomotor activity, has anxiolytic and antidepressant properties, and corrects sensorimotor and motor coordination impairments in animal models. Suggested mechanisms involve the modulation of neurotransmitter systems, interference with pro-inflammatory cytokine production, reduction of oxidative neuronal damage, and activation of neurotrophic signaling pathways [33]. Rats with temporal lobe epilepsy based on the kainic acid model were employed in a 2019 study to evaluate the impact of apigenin. Apigenin was given orally at 50 mg/kg for six days. The delayed onset and reduced severity of seizures reflected significant anticonvulsant activity. Apigenin also promoted survival in hippocampus hilus neurons and enhanced memory impairments induced by epilepsy. Apigenin decreased cytochrome c release, as indicated by immunohistochemical evaluation, which would indicate that it prevents the mitochondrial apoptotic pathway, one of which may be its neuroprotective effects [34]. Research on vitexin, designated as apigenin-8-C-glucoside, a flavonoid with a structural similarity to apigenin. Vitexin was delivered intraperitoneally at doses of 1.25 to 5 mg/kg in murine models. The research concluded that vitexin exerted dose-dependent protection against seizures induced by GABAergic antagonists like picrotoxin and pentylenetetrazol (PTZ), with the maximum dose conferring protection against tonic-clonic seizures. Nonetheless, vitexin was ineffective against seizures induced by glutamate receptor agonists such as NMDA and kainic acid. Vitexin was also found to have anxiolytic-like activity with no sedation or locomotor impairment, indicating its possible role as a modulator of GABAergic neurotransmission [35]. Another study in 2020 explored apigenin’s impact on oxidative stress and ferroptosis in epileptic brains. The study revealed that apigenin was able to reduce myeloperoxidase-induced oxidative stress and block ferroptosis, a type of regulated cell death in epilepsy. Table 1 presents the key pharmacological activities of apigenin along with their associated molecular pathways, potential drug partners, and proposed synergistic roles. The observations point towards the involvement of the antioxidant activity of apigenin in its anticonvulsant action. Collectively, these findings highlight apigenin’s potential as a neuroprotective adjuvant. Its modulation of oxidative stress and neuroinflammatory pathways supports its synergistic application with existing neurotherapeutics to enhance cognitive function and neuronal survival [36].
Apigenin activities, associated molecular pathways, potential drug partners, and reported synergistic roles supported.
| Activity | Key pathways | Potential drug partners | Synergistic role |
|---|---|---|---|
| Anticancer [37] | PI3K/Akt, NF-κB, p53 | Doxorubicin, cisplatin, TRAIL | Enhances apoptosis, reduces resistance |
| Anti-inflammatory [38] | COX-2, NF-κB, cytokines | NSAIDs, corticosteroids | Amplifies anti-inflammatory effects |
| Antioxidant [39] | Nrf2, ROS modulation | Chemotherapeutics | Reduces oxidative stress-mediated toxicity |
| Antidiabetic [40] | AMPK, GLUT4 | Metformin | Improves insulin sensitivity |
| Antibacterial [41] | Membrane disruption, quorum sensing | Antibiotics | Enhances drug uptake, reduces resistance |
The study tested the antibacterial properties of certain flavonoids like apigenin against several bacterial strains. The results showed that apigenin only had weak inhibitory effects on Staphylococcus aureus and no apparent antibacterial effect on other tested bacterial species. This suggests that apigenin may act on bacteria but is not a good antibacterial agent, and more studies are needed to know how it acts and whether it has synergism with other compounds [42]. The study examined the apoptosis-like antibacterial activity of apigenin against Escherichia coli. It was found that apigenin causes bacterial cell death by causing an increase in intracellular calcium levels. Elevated levels of calcium trigger the production of reactive nitrogen and oxygen species that cause oxidative stress. The oxidative stress causes damage to the bacterial cell membrane, leading to cell disruption and death. This action highlights the antimicrobial activity of apigenin through its ability to produce oxidative injury in bacterial cells [43]. Recent research examined the anti-biofilm activity of apigenin-7-O-glucoside (A7G) against Staphylococcus aureus and Escherichia coli. The research demonstrated that A7G significantly inhibits biofilm formation through the inhibition of exopolysaccharide production, a requirement for biofilm stability, disruption of quorum-sensing mechanisms governing bacterial communication, and reducing cell surface hydrophobicity, which is involved in bacterial adherence. These findings suggest that A7G has high potential as a natural anti-biofilm and antibacterial molecule, which could be useful in the elimination of biofilm-producing bacteria that cause chronic infections [44]. The antipathogenic ability of apigenin was recently studied in its ability to combat Helicobacter pylori, a bacterium linked to gastritis and peptic ulcers. Researchers developed a gastroprotective microsponge formulation of apigenin that showed potent in vitro antibacterial activity against a metronidazole-resistant Helicobacter pylori strain. The microsponge formulation worked especially well with antibacterial action that was maintained for up to 72 hours, compared to 36 hours when using pure apigenin. This sustained effect underscores the formulation’s therapeutic potential for long-term action, constituting a promising vehicle for the treatment of Helicobacter pylori infections, especially those refractory to conventional therapies. Despite its moderate standalone antibacterial activity, apigenin demonstrates significant potential for synergistic interactions. Its ability to disrupt bacterial membranes and induce oxidative stress may enhance antibiotic efficacy and help combat antimicrobial resistance [45].
In an experimental study, the apigenin effects were investigated in a murine model of asthma with mice that were sensitized and ovalbumin (OVA) challenged to induce a model of allergic asthma response. When apigenin-treated mice, the asthma symptoms significantly improved. Apigenin reduced inflammatory cell invasion into the lungs, which is typically a feature of inflammation in asthma [46]. It also inhibited airway hyperresponsiveness, an over-reaction of the airway to stimuli, and decreased total immunoglobulin E (IgE) levels, a key antibody in allergy. Apart from that, apigenin altered the immune response from the T-helper type 2 (Th2) predominant pattern, typical of allergic inflammation, to the pattern of a T-helper type 1 (Th1) profile. Th1 responses are also accompanied by anti-inflammatory actions and a reduced tendency for allergic reactions. This conversion shows that apigenin may possess anti-inflammatory activities that can benefit in managing allergic asthma by modulating immune reactions [47]. The research investigated the anti-asthmatic effects of apigenin in an OVA-induced asthma model. Apigenin treatment caused decreased airway resistance, decreased numbers of eosinophils, and decreased pro-inflammatory cytokine levels (IL-6, TNF-α, and IL-17A). The research also detected strong inhibition of T-helper 17 (Th17) cells and down-regulation of RORγt protein expression, suggesting that apigenin could be useful in managing asthma progression through modulation of the Th17 cell response and reduction of inflammation [48]. Research compared the actions of natural flavones, such as apigenin, on late-phase and immediate asthmatic responses in guinea pigs sensitized to OVA. Apigenin therapy significantly reduced certain airway resistance in both phases and inhibited leukocyte mobilization, histamine release, and activity of phospholipase A2 and eosinophil peroxidase in bronchoalveolar lavage fluid. These findings show that apigenin possesses anti-asthmatic activity through inhibition of airway hyperresponsiveness and inflammation [49]. An experimental study investigated apigenin’s potential to alleviate airway inflammation in PM2.5-exposed asthmatic mice. Apigenin significantly decreased airway hyperresponsiveness and suppressed eosinophil and neutrophil infiltration in bronchoalveolar lavage fluid and lung tissue. The study also showed significant reductions in total serum IgE and cytokines IL-4, IL-13, and IL-17 levels. In addition, apigenin suppressed the mRNA level of the NF-κB p65 subunit in lung tissue, which implies its involvement in inflammatory response modulation in asthma induced by PM2.5 exposure. These immunomodulatory and anti-inflammatory effects suggest that apigenin may act synergistically with conventional anti-asthmatic drugs. Such combinations could improve therapeutic outcomes by targeting multiple inflammatory pathways simultaneously [50].
Researchers isolated apigenin from the Portulaca Oleracea plant and evaluated its antibacterial activity against several pathogenic bacteria, including Salmonella typhimurium. The study reported a zone of inhibition of 17.36 ± 0.18 mm for Salmonella typhimurium, indicating significant antibacterial activity. However, the minimum inhibitory concentration (MIC) for all tested bacterial strains, including Salmonella typhimurium, was found to be greater than 4 mg/mL, suggesting that higher concentrations of apigenin are required to inhibit bacterial growth effectively [51]. This study focused on the antibacterial mechanism of apigenin in Escherichia coli. The researchers found that apigenin induced apoptosis-like death in Escherichia coli by increasing intracellular calcium levels and promoting the production of reactive nitrogen and oxygen species. While this study did not directly assess apigenin’s effect on Salmonella typhimurium, the findings suggest a potential mechanism by which apigenin could exert antibacterial effects on similar Gram-negative bacteria [43]. This study examined the inhibitory activity of apigenin against multiple bacterial species. The results for Salmonella typhimurium were mixed, with some studies reporting no significant inhibition, while others observed antibacterial effects. These discrepancies may be attributed to differences in experimental conditions, bacterial strains, and apigenin concentrations used. A comprehensive review highlighted apigenin’s antimicrobial potential, noting that its inhibitory effects are strain-specific. In certain cases, although apigenin did not inhibit the growth of pathogenic bacteria, it reduced toxin production and mitigated pathogen-induced injury. Additionally, synergistic effects have been observed when apigenin is combined with other antibiotics. Although apigenin requires higher concentrations for direct antibacterial effects, its mechanism of inducing oxidative stress in bacteria supports its potential as a synergistic agent. Combination with standard antibiotics may enhance efficacy against resistant strains [52].
Researchers demonstrated that apigenin inhibits the expression of adhesion molecules induced by high glucose and tumor necrosis factor-alpha in human endothelial cells, suggesting a protective role against vascular complications in diabetes [53]. They found that apigenin improves diabetes by modulating biochemical pathways, enhancing GLUT4 translocation, and altering CD38 expression, thereby improving glucose uptake and metabolism [54]. A study by Ren et al. showed that apigenin ameliorates glucose and lipid metabolism disorders and improves vascular dysfunction in type 2 diabetic rats, indicating its multifaceted benefits in managing diabetes [55]. A systematic review highlighted apigenin’s role in alleviating insulin resistance, regulating glycolipid metabolism, and mitigating oxidative stress through various mechanisms, including the inhibition of insulin receptor kinase activity and modulation of microRNAs [23]. Recent research focused on synthesized apigenin analogs revealed significant inhibitory effects on α-glucosidase, an enzyme involved in carbohydrate digestion, suggesting potential therapeutic applications for type 2 diabetes. A study published in the Uttar Pradesh Journal of Zoology examined the effects of apigenin in alloxan-induced diabetic rats. The outcomes reported that apigenin dramatically lowered blood glucose levels, suppressed the activity of α-amylase, and improved the activity of antioxidant enzymes. The outcomes indicate that apigenin alone and combined with metformin could be efficacious in the management of hyperglycemia and oxidative stress in diabetes [56]. Table 2 provides a consolidated overview of the diverse pharmacological activities of apigenin, along with their proposed mechanisms and experimental models. A published study in the European Journal of Pharmacology reviewed the impact of apigenin on cognitive impairment caused by hyperglycemia in zebrafish. The test revealed that apigenin improved learning and spatial memory, reduced blood glucose levels, and mitigated brain oxidative stress. Such protective effects on the brain were attributed to Nrf2/ARE pathway induction, highlighting apigenin’s potential in reducing diabetic-related cognitive impairment.[57]. Research work in Bioorganic Chemistry examined synthesized apigenin analogs as α-glucosidase inhibitors. Among the compounds being tested, one analog was reported to possess significant inhibitory activity, lowering fasting blood glucose and reducing insulin resistance in a mouse model of type 2 diabetes. This points to the potential of apigenin derivatives in the development of new antidiabetic agents [58]. Apigenin has been studied for its influence on pancreatic β-cells and has been reported to increase glucose-stimulated insulin secretion together with protection against ER stress-induced apoptosis. Such protective effects may be attributed to the possible downregulation of proteins crucial for stress, namely, CHOP and TXNIP, by apigenin. Modulation of these molecular pathways helps apigenin to preserve β-cell function and cell survival, which could suggest a therapeutic role for this flavonoid in preventing or managing diabetes-related β-cell dysfunction. These findings indicate that apigenin holds strong potential as an adjunct in diabetes management. Its ability to regulate glucose metabolism and oxidative stress supports synergistic use with antidiabetic drugs such as metformin to improve glycemic control and reduce complications [59].
Summary of apigenin’s biological effects observed in vitro, in vivo, and clinical studies, highlighting key outcomes, mechanisms, and translational relevance.
| Methods | Typical experimental system/dose ranges | Main effects observed | Putative mechanisms | Strength |
|---|---|---|---|---|
| In vitro (cells, biochemical assays) [60] | Cancer /immune/neuronal/endothelial cell lines; typical concentrations ~1–100 µM (commonly 5–50 µM) |
| Modulation of signalling (PI3K/Akt, MAPK, NF-κB), upregulation of pro-apoptotic proteins, inhibition of kinases and MMPs, and antioxidant enzyme activation. | Strong, consistent mechanistic data across many cell types, but limited to controlled cell conditions (not directly predictive of human dosing). |
| In vivo (animal models) [61] | Rodent disease models; doses often ~1–200 mg/kg (oral or i.p.) depending on model and duration |
| Same pathways as in vitro plus effects on whole-organism pharmacology (reduced systemic cytokines, oxidative stress; improved organ histology). Often shows chemosensitization when combined with drugs. | Good preclinical efficacy and safety signals, but results vary by formulation, dose, and model—translation to humans uncertain. |
| Clinical/human studies [62] | Very limited controlled trials and PK studies; human trials mostly early-phase or pilot; clinicaltrials.gov lists PK/safety and small pilot studies |
| Human metabolism (glucuronidation/sulfation) strongly affects exposure; formulations and glycoside forms change absorption. | Insufficient clinical efficacy data. Safety in short studies appears acceptable, but robust efficacy and dosing regimens are not established—more human trials needed. |
The combination of apigenin with conventional chemotherapeutic agents has emerged as a promising strategy to enhance anticancer efficacy while overcoming drug resistance. Apigenin has been shown to potentiate the therapeutic effects of drugs such as doxorubicin, cisplatin, and TRAIL through multi-targeted mechanisms involving apoptosis induction, oxidative stress modulation, and regulation of key signaling pathways [63]. The co-administration of apigenin with doxorubicin has demonstrated significant enhancement in anticancer activity across various cancer cell lines. This combination leads to increased intracellular accumulation of doxorubicin, primarily through the inhibition of efflux transporters such as P-glycoprotein, thereby reducing multidrug resistance. Mechanistically, apigenin promotes p53 activation, enhances ROS generation, and facilitates mitochondrial-mediated apoptosis. The combined treatment results in amplified DNA damage, caspase activation, and a marked increase in apoptotic cell death compared to monotherapy [64].
Similarly, the synergistic interaction between apigenin and cisplatin has been widely reported to improve therapeutic outcomes. Apigenin sensitizes cancer cells to cisplatin by modulating key survival pathways such as PI3K/Akt and NF-κB, which are often implicated in chemoresistance. Additionally, apigenin enhances oxidative stress within tumor cells, leading to mitochondrial dysfunction and apoptosis [65]. This combination has been shown to reduce cisplatin-induced toxicity while maintaining or enhancing anticancer efficacy, thereby improving the therapeutic index. Apigenin also exhibits strong synergy with TRAIL, particularly in TRAIL-resistant cancer cells [66]. The combination enhances apoptosis by upregulating death receptors DR4 and DR5 in a p53-dependent manner, thereby restoring TRAIL sensitivity. Furthermore, apigenin inhibits anti-apoptotic proteins such as Bcl-2 and survivin, while activating cascades. Collectively, these interactions result in increased apoptotic signaling and reduced tumor cell survival, highlighting apigenin’s potential as a sensitizing agent in targeted cancer therapies [67].
Apigenin has demonstrated promising synergistic effects when combined with standard antidiabetic agents such as metformin, offering improved glycemic control and metabolic regulation. Metformin is a first-line oral antihyperglycemic agent widely used in the management of type 2 diabetes mellitus and acts primarily by reducing hepatic glucose production and improving insulin sensitivity [68]. The combination of apigenin with metformin has been shown to enhance glucose uptake and utilization through modulation of GLUT4 translocation and activation of AMPK pathways. Apigenin complements metformin’s action by improving insulin sensitivity and reducing insulin resistance at the cellular level. Additionally, apigenin exerts antioxidant effects that mitigate oxidative stress, a key contributor to diabetic complications [69].
Recent studies also suggest that metformin may exhibit indirect or adjuvant antimicrobial effects, possibly mediated through host metabolic modulation, alteration of gut microbiota, and AMPK-dependent pathways. While metformin is not classified as an antibiotic, these findings highlight its potential ancillary role in influencing microbial processes [70]. Mechanistically, apigenin regulates key enzymes involved in carbohydrate metabolism and suppresses inflammatory mediators associated with insulin resistance. The combined treatment has been reported to improve lipid profiles, reduce hyperglycemia, and protect against vascular complications. This synergistic interaction suggests that apigenin can act as an effective adjuvant to conventional antidiabetic therapy, enhancing therapeutic outcomes while potentially lowering drug dosage requirements [71].
The combination of apigenin with conventional antibiotics represents a promising strategy to combat antimicrobial resistance and improve therapeutic efficacy. Although apigenin alone exhibits moderate antibacterial activity, its synergistic interactions with antibiotics significantly enhance antimicrobial effects against a range of pathogenic microorganisms. Apigenin has been shown to disrupt bacterial cell membrane integrity, leading to increased permeability and facilitating enhanced antibiotic uptake [72]. This membrane-disruptive action weakens bacterial defense mechanisms and potentiates the activity of co-administered antibiotics. Additionally, apigenin interferes with bacterial quorum-sensing systems, thereby inhibiting communication pathways essential for virulence and survival. A key mechanism underlying this synergy is the inhibition of biofilm formation, which is a major factor contributing to chronic infections and antibiotic resistance. Apigenin reduces exopolysaccharide production and alters cell-surface properties, thereby preventing bacterial adhesion and biofilm maturation. These effects enhance bacterial susceptibility to antibiotics and improve treatment outcomes. Furthermore, apigenin-induced oxidative stress within bacterial cells contributes to cellular damage and apoptosis-like death, further amplifying antimicrobial efficacy. This combination approach holds significant potential in addressing multidrug-resistant infections and reducing the burden of antibiotic resistance [73].
Emerging evidence suggests that apigenin may enhance the efficacy of neuroprotective agents through synergistic interactions targeting oxidative stress, neuroinflammation, and neuronal survival pathways. Although studies in this area are relatively limited, available data indicate promising therapeutic potential in neurodegenerative disorders. Apigenin, when used in combination with neuroprotective drugs or agents targeting cognitive impairment, has been shown to improve neuronal function and reduce neurotoxicity. The synergistic effects are primarily mediated through the modulation of PI3K/Akt, ERK/CREB/BDNF, and Nrf2 signaling pathways, which play critical roles in neuronal survival, synaptic plasticity, and antioxidant defense [74]. Additionally, apigenin enhances mitochondrial function, reduces the accumulation of neurotoxic proteins such as amyloid-β, and attenuates neuroinflammation by inhibiting pro-inflammatory cytokines. These combined effects contribute to improved cognitive performance and neuroprotection in experimental models. The integration of apigenin with existing neurotherapeutics may offer a multi-targeted approach for the management of neurodegenerative diseases such as AD and epilepsy. However, further studies are required to validate these findings in clinical settings and to establish optimal dosing strategies for combination therapy [75]. Table 3 summarizes key studies demonstrating the synergistic effects of apigenin in combination with conventional small-molecule drugs, highlighting therapeutic outcomes, underlying mechanisms, and experimental models.
Summary of synergistic interactions between apigenin and small-molecule drugs
| Drug | Disease/model | Effect of combination with apigenin | Proposed mechanism of synergy | Study type | Quantitative synergy metric |
|---|---|---|---|---|---|
| Doxorubicin [76] | Breast/various cancers | Enhanced apoptosis and reduced drug resistance | ROS generation, p53 activation, cell cycle arrest | In vitro | CI = 0.65–0.85 (reported) |
| Cisplatin | Ovarian/lung cancer | Increased cytotoxicity and chemosensitivity | Inhibition of the PI3K/Akt pathway, apoptosis induction | In vitro/in vivo | CI < 1 (qualitative report) |
| TRAIL [11] | Lung cancer | Increased sensitivity to apoptosis | Upregulation of DR4/DR5 receptors (p53-dependent) | In vitro | Not reported |
| Metformin [56] | Type 2 diabetes | Improved glucose regulation and insulin sensitivity | Activation of AMPK, enhanced GLUT4 translocation | In vivo | Not applicable |
| 5-Fluorouracil [76] | Colorectal cancer | Enhanced anticancer efficacy | NF-κB inhibition, apoptosis enhancement | In vitro | CI ~0.7 (reported) |
| Paclitaxel [77] | Breast cancer | Increased tumor cell death | Microtubule stabilization + apoptosis pathways | In vitro | Synergistic (CI < 1; exact value not reported) |
| Antibiotics (e.g., Ciprofloxacin) [78] | Bacterial infections | Enhanced antibacterial activity, reduced resistance | Membrane disruption, biofilm inhibition | In vitro | Not reported |
| Temozolomide [79] | Glioblastoma | Increased chemosensitivity | ROS generation, DNA damage enhancement | In vitro | Synergistic (qualitative; CI not reported) |
CI (Combination Index) values are based on the Chou–Talalay method where reported. CI < 1 indicates synergism, CI = 1 indicates an additive effect, and CI > 1 indicates antagonism. “Not reported” denotes that quantitative synergy metrics were not provided in the original study.
Some studies have investigated the synergistic effects of apigenin when used with other therapeutic agents. Apigenin markedly enhanced the anticancer efficacy of 5-fluorouracil in ErbB2-overexpressing MDA-MB-453 cells. In comparison with single 5-FU treatment, the combination induced more significant growth inhibition and apoptosis, possibly through suppression of ErbB2 expression and inhibition of the Akt signaling pathway [80]. In MDA-MB-231 TNBC cells, apigenin in combination with the histone deacetylase inhibitor vorinostat (SAHA) triggered severe apoptotic cell death, such as nuclear fragmentation, chromatin condensation, and low mitochondrial membrane potential. These data indicate that apigenin can augment the therapeutic effect of vorinostat by modulating both epigenetic and apoptotic regulators [81]. Besides, apigenin was found to be synergistic with doxorubicin in estrogen-dependent (MCF-7) and non-estrogen-dependent (MDA-MB-231) breast cancer cells. It was found to be dose-dependent for cytotoxicity and anti-migration activities, and both drugs induced lipid droplet accumulation in MDA-MB-231 cells, which may be related to interaction with ATP-binding cassette transporters and modulation of AKT and MYC pathways [76]. Besides cancer therapy, apigenin has also shown promise in infectious disease management. When combined with β-lactam antibiotics ampicillin and ceftriaxone, apigenin significantly improved their action against methicillin-resistant Staphylococcus aureus (MRSA), suggesting its ability to counter antibiotic resistance [82].
The therapeutic potential of apigenin in metabolic wellness has been clarified by recent studies, particularly when combined with other bioactive substances. A study examined the effects of apigenin and resveratrol on the trans differentiation and plasticity of white adipocytes [83]. When compared to each drug alone, the combination of treatments was more successful in browning white adipocytes. This augmented effect was attributed to the activation of more than one signaling pathway, such as PI3K signaling through an estrogen receptor-dependent pathway, ultimately enhancing angiogenesis and lipid metabolism [84]. These observations project the combination of apigenin and resveratrol as a promising treatment to enhance lipid metabolism in obese subjects [85]. Another study investigated the interaction of apigenin with emodin, naringin, and quercetin in suppressing 3T3-L1 preadipocyte differentiation and pancreatic lipase activity. In the combinations examined, emodin and apigenin exhibited the most significant synergistic effects in both assays and corroborate their complementary therapy role for obesity control [86]. This implies that apigenin can act against obesity partially through prebiotic-like modulation of the gut microbiome, which further attests to its promise in metabolic health interventions [87]. Apigenin has been shown to have potential therapeutic benefits in the field of metabolic health, particularly if combined with other bioactive compounds.
An experiment tested the combination of emodin, naringin, and quercetin in inhibiting 3T3-L1 preadipocyte differentiation and pancreatic lipase activity. Its value as an adjunct drug in controlling overweight and obesity is evidenced by the fact that emodin-apigenin showed the highest synergistic effect among all the combinations used in both assays. Especially when co-administered with other bioactive compounds, apigenin has shown promising therapeutic potential in the context of metabolic health [86]. Despite these encouraging results, the authors indicated the need for more studies to establish the mechanism of action and clinical uses [88]. In the same vein, a systematic review investigated the role of apigenin in the targeting of metabolic syndrome and emphasized its antiobesity properties, antidiabetic activity, and ability to improve metabolic dysfunction [89]. The review stated that apigenin is a very promising drug as a therapeutic agent and further called for more human clinical trials to support its efficacy and safety in the management of metabolic syndrome [90].
While apigenin is generally known for its medicinal promise, attention must be given to its safety profile, especially at toxic doses or as a supplement [91]. Genotoxicity testing has indicated that apigenin, at high levels, can present genotoxic activity in some test systems, and there are fears about the genetic risks of overdose [92]. Sedative effects have also been observed in animal studies, where high doses (100 and 200 mg/kg) led to mild sedation and muscle relaxation, suggesting central nervous system impacts that could impair alertness or induce drowsiness [93]. In addition to its antioxidant activity, apigenin has been reported to interfere with hormone-related pathways, particularly those involving estrogen and testosterone. Inhibition of these enzymes will change the blood levels of some medications to either decreased effect or increased risk of toxicity [94]. Elevated doses of apigenin have also been linked to gastrointestinal upset, such as stomach discomfort, nausea, and diarrhea. People with an allergy to plants, especially those who are allergic to plants in the Asteraceae group (e.g., chamomile or daisies), are also likely to suffer from an allergic reaction that could be anything from skin rashes to respiratory distress [95]. According to the overall safety, a systematic review has found that although apigenin is well-tolerated when ingested via dietary intake, caution should be practiced with high-dose supplementation because of the risk of side effects like sedation, hormonal activity, and drug interactions. The review firmly recommends additional clinical studies to determine definite and safe dosages [96]. In addition, liver toxicity has also been documented in animal models, where the doses of approximately 100 mg/kg resulted in manifestations of liver damage, although human evidence is still sparse. Generally, though apigenin is a promising drug, its use in a safe manner requires cautious regard for dosage, the health status of the individual, and interactions [97]. The mechanistic pathways and pharmacokinetic profile of apigenin have been discussed earlier (Figure 4).
To maximally realize the therapeutic value of apigenin, several strategic avenues are being pursued. One is the increase of its bioavailability using new drug delivery systems, including nanocarriers and liposomes, designed to enhance its solubility and stability. At the same time, properly conducted human clinical trials are needed to confirm apigenin’s efficacy and safety in various health conditions [98]. It is also important to understand more precise mechanistic facts, such as how it works at the molecular level and the cellular pathways it acts on. Furthermore, apigenin’s synergistic action when used with other natural compounds or drugs is being investigated increasingly, which can help develop more potent treatments. Apart from its medicinal application, apigenin is rich with potential applications in industry as a pharmaceutical excipient, dietary supplement, and functional food component, further enhancing its influence on overall health and disease prevention [99].
Apigenin is a multifunctional dietary flavonoid with broad pharmacological activities, including anticancer, anti-inflammatory, antioxidant, neuroprotective, and metabolic regulatory effects. Beyond its standalone efficacy, accumulating evidence underscores its role as a potent bioenhancer in combination therapy. Notably, apigenin exhibits strong synergistic interactions with chemotherapeutic agents such as doxorubicin, cisplatin, paclitaxel, and TRAIL, where it enhances apoptosis, suppresses drug resistance through P-glycoprotein inhibition, and modulates key signaling pathways including PI3K/Akt and NF-κB. Similarly, its combination with antidiabetic agents like metformin improves glycemic control via AMPK activation and enhanced GLUT4 translocation, while antimicrobial synergy with antibiotics contributes to overcoming drug resistance through membrane disruption, biofilm inhibition, and increased drug uptake. These multi-targeted interactions highlight apigenin’s potential as an adjuvant therapeutic agent capable of improving efficacy while reducing toxicity and dosage requirements of conventional drugs. However, despite promising preclinical evidence, the translational application of these synergistic combinations remains limited due to poor bioavailability, lack of standardized dosing strategies, and insufficient clinical validation. Future research should prioritize well-designed clinical trials, pharmacokinetic optimization, and quantitative synergy modeling to establish the safety, efficacy, and therapeutic relevance of these combinations in humans. Overall, apigenin-based combination strategies represent a promising avenue for next-generation therapeutics, bridging natural product pharmacology with modern precision medicine.
AD: Alzheimer’s disease
HIF-1α: hypoxia-inducible factor-1 alpha
IgE: immunoglobulin E
OVA: ovalbumin
PC: pancreatic cancer
ROS: reactive oxygen species
Th2: T-helper type 2
The authors express their sincere gratitude to their respective institutions, including IFTM University, Moradabad; Raj Kumar Goel Institute of Technology (Pharmacy), Ghaziabad, SRM Institute of Science and Technology, Delhi-NCR Campus; Moradabad Educational Trust, Group of Institutions; Saveetha Institute of Medical and Technical Sciences, Chennai; and Chitkara University, Punjab, for providing the necessary academic support and research environment to carry out this work. The authors acknowledge the use of ChemDraw (PerkinElmer Informatics) for generating chemical structure illustrations.
AK: Conceptualization, Methodology, Writing—review & editing, Supervision. HS: Methodology, Investigation. RV: Investigation, Data curation. SK: Methodology, Investigation. JA: Formal analysis, Data curation, Writing—original draft, Visualization. PK: Formal analysis, Writing—original draft. AKM: Resources, Writing—original draft, Visualization. SC: Validation, Resources, Writing—review & editing. HC: Conceptualization, Validation, Writing—review & editing, Supervision. All authors have read and agreed to the published version of the manuscript.
The authors declare that they have no competing interests.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
The authors received no specific funding for this work.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 795
Download: 11
Times Cited: 0