 Review
Review

Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
†These authors contributed equally to this work.
Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
†These authors contributed equally to this work.
Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
Email: vrai@westernu.edu
ORCID: https://orcid.org/0000-0001-6286-2341
Explor Drug Sci. 2026;4:1008168 DOI: https://doi.org/10.37349/eds.2026.1008168
Received: March 04, 2026 Accepted: May 18, 2026 Published: July 15, 2026
Academic Editor: Juergen Reichardt, James Cook University, Australia
The article belongs to the special issue Advances and Innovations in Gene-Based Medicine
A diabetic foot ulcer (DFU) is an open sore or wound, usually on the bottom of the foot, affecting about 15% of people with diabetes. Caused by nerve damage, poor circulation, and high pressure, these ulcers often present as swelling, drainage, redness, or foot deformities. Treatment focuses on infection control, debridement, and offloading pressure, often taking weeks to months to heal. Despite treatment, recurrence and amputation remain major clinical concerns. Gene therapy for DFUs is an emerging, promising field aiming to accelerate healing in chronic, non-healing wounds by delivering therapeutics (e.g., growth factors) directly to the site to promote angiogenesis and tissue regeneration. Key approaches include topically applied, multi-target gene therapies (e.g., AUP1602-C reporting 83% healing rate) and plasmid-based therapies (e.g., VM202/Engensis). This narrative review aims to describe the molecular aspects of wound healing, pathogenesis and management of DFU, followed by evolving gene therapies including AUP-16, VM202, and gene targets like SCUBE1, RNF103-CHMP3 and PGI1. We have also discussed emerging targets such as C-X-C motif chemokine receptor 4 (CXCR4), thrombospondin 1 (TSP1), yes-associated protein-transcriptional co-activator with PDZ binding motif (YAP-TAZ) pathway, and others as potential targets for gene therapy to promote healing in chronic non-healing DFUs.
Diabetes mellitus (DM) is a chronic metabolic condition characterized by chronic hyperglycemia that, as of 2021, affects roughly 11.6% of the United States population. Various complications can arise from improper treatment and management of DM, most notably diabetic foot ulcers (DFUs). According to the International Diabetes Federation (IDF) Diabetes Atlas, approximately 6.3% of patients with DM worldwide are affected by DFUs, and 19–34% of diabetic patients develop a DFU in their lifetime. 20% of which will require an amputation at some point, and 10% of patients diagnosed with a DFU will die within 1 year of their diagnosis [1].
DFUs primarily develop because of chronic hyperglycemia, which leads to major pathological processes: diabetic neuropathy (DN) and peripheral artery disease (PAD) [2]. DN manifests in three forms: sensory, motor, and autonomic neuropathy. Sensory neuropathy involves the loss of protective sensation in the extremities, diminishing pain, temperature, and pressure perception. As a result, minor wounds and sores go unnoticed and untreated. Motor neuropathy results from the denervation of intrinsic foot muscles, leading to muscle imbalance, foot deformities, and abnormal plantar pressure distribution. These structural changes increase mechanical stress and predispose the skin to breakdown. Ill-fitting footwear exacerbates this issue by creating friction and pressure points that increase the likelihood of wound formation. Autonomic neuropathy impairs the regulation of sweat and oil glands, resulting in dry, cracked skin that compromises the skin barrier [3]. While minor lesions may heal under normal physiological conditions, in diabetic patients who have PAD, the impaired blood flow to the affected area results in tissue ischemia. Reduced delivery of oxygen, immune cells, and nutrients to the affected site compromises wound healing, reduces the ability to combat infection, and disrupts the maintenance of healthy skin and nerves [4].
The high glucose microenvironment also disrupts normal cell function, which impairs the immune response, delaying healing. Chronic inflammation of the site disrupts growth factor signaling, inhibiting progression of the normal phases of wound healing. The elevated presence of glucose also promotes bacterial growth and biofilm formation. Once bacteria invade the system, infection can set in and result in further complications [5]. Despite adequate management, DFUs remain vulnerable to microbial infections and are susceptible to reinfection. Nearly 50% of patients experience reinfection, and up to 65% face recurrence within 5 years. High-risk factors include long-lasting wounds, bone infection (osteomyelitis), poor glycemic control, PAD, and, significantly, the presence of bacterial biofilms that impair skin barrier function. This contributes to chronic non-healing DFUs due to a persistent inflammation response (chronic inflammation) that is far more intense compared to a regular acute response during wound healing [5, 6].
Current treatments, including conventional strategies, oxygen therapy, stem cell therapy, platelet-rich plasma therapy, and skin grafting, have shown promising results but are in the developing stage. However, reinfection, recurrence, and amputation in DFUs indicate the need for new therapeutic approaches. This is important because patients undergoing treatment, even with early stages of DFU, still have a relatively high chance of failed treatment leading to undesirable patient outcomes [7]. New treatments or care strategies must target the underlying pathophysiology rather than merely treating symptoms. Recent reports investigating the underlying molecular mechanisms [8–11] have reported the role of differentially expressed genes, proteins, and microRNAs (miRNAs) in non-healing diabetic wounds compared to nondiabetic wounds or normal skin. This suggests that differential expression of a gene may be a potential therapeutic target. By targeting specific genes playing a pathophysiologic role in DFU formation and non-healing, gene expression could be regulated to promote healing and maximize patient outcomes. This narrative review aims to discuss normal wound healing, wound healing in diabetics, the pathophysiology, current therapeutics, and treatment aspects of DFUs, followed by a summary of the emerging gene-based therapies and emerging therapeutic targets for DFUs. Rather than addressing clinical manifestations, these approaches aim to treat underlying molecular and cellular dysfunctions that drive non-healing wounds.
The normal wound healing process occurs through four overlapping stages: hemostasis, inflammation, proliferation, and remodeling. The hemostasis stage is responsible for rapidly controlling bleeding and re-establishing the physical barrier of the epidermis. This phase is initiated by vasoconstriction, followed by the aggregation of platelets and fibrin. This together forms a scaffold (platelet plug) that prevents bacteria from entering through the exposed wound and limits blood loss [12] (Figure 1). In patients with diabetes, the normal stages of wound healing are significantly impaired because of PAD and hyperglycemia. During the hemostasis phase, chronic hyperglycemia promotes the formation of advanced glycation end products (AGEs), which accumulate on extracellular matrix (ECM) proteins. This accumulation disrupts normal cell matrix interactions and signaling, resulting in a dysfunctional wound matrix and delaying the next stages of wound healing [13] (Figure 1).
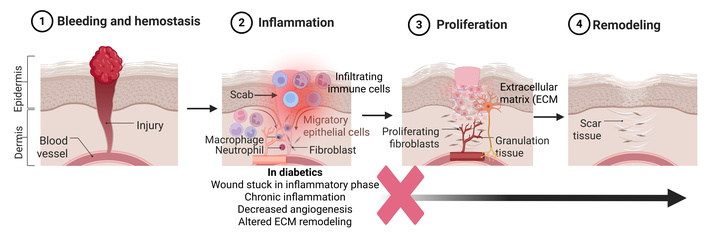
Wound healing and changes during diabetes in diabetic wound healing. The figure shows normal healing and the events in these phases. In the case of chronic ulcers and diabetic foot ulcers, wounds get stuck in the inflammatory phase and do not progress to the proliferation phase (marked by a red cross sign). The black arrow shows that the progression of the wound in the proliferation and remodeling phase, which normally occurs during wound healing, does not happen in chronic diabetic wounds. Created in BioRender. Rai, V. (2026) https://BioRender.com/8g8wbna.
In the second stage, known as the inflammation stage, it is essential for wound decontamination and early tissue repair. Here, neutrophils and macrophages secrete inflammatory factors to induce vasodilation to increase the amount of oxygen and leukocytes to the affected area. Helping to eliminate pathogens and remove cellular debris, preparing the wound bed for subsequent tissue regeneration. Later on in the inflammation stage, the macrophages change from a proinflammatory to an anti-inflammatory phenotype [12, 14] (Figure 1). The inflammatory phase in diabetic wounds is both prolonged and ineffective. Immune cells exhibit impaired pathogen clearance, impaired leukocyte chemotaxis, phagocytosis, and bactericidal activity, resulting in sustained inflammatory signaling due to chronic hyperglycemia. Macrophage dysfunction further exacerbates impaired healing, as diabetic wounds fail to undergo the normal transition of macrophages from the proinflammatory (M1) phenotype to the reparative (M2) phenotype required for angiogenesis and tissue regeneration. Pro-inflammatory cytokines, including interleukin (IL)-6, IL-1β, IL-8, tumor necrosis factor (TNF)-α, and others, remain elevated, maintaining the wound in a chronic inflammatory state (Figure 1). This persistent inflammation drives excessive protease activity and disrupts growth factor signaling, inhibiting progression to the proliferative phase of wound healing [15] (Figure 1). Most of the diabetic wounds are stuck in this stage due to persistent inflammation.
Simultaneously, the proliferation stage rebuilds tissue as fibroblasts deposit collagen and glycosaminoglycans. Replacing the fibrin from the hemostasis stage and stabilizes the wound. Re-epithelialization starts to occur once keratinocytes begin to migrate from the periphery of the wound to form new layers of epithelial cells that gradually thicken over time. At the same time, the dermal layer contracts to help seal the wound. Granulation tissue fills the wound. Then, the formation of new blood vessels through angiogenesis and vasculogenesis restores blood flow and oxygen delivery to the previously damaged area [14, 16]. In diabetic wounds, however, angiogenesis is significantly impaired due to reduced expression of key growth factors such as vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF)-2. VEGF initiates angiogenesis and mediates endothelial cell proliferation, while FGF-2 facilitates migration of new blood vessels through the ECM [17]. If the expression of these factors is reduced, the wound’s capability to heal is reduced. In addition, in patients with type 2 diabetes, the accumulation of AGEs induces senescence and contributes to both a reduction in the number and impaired function of endothelial progenitor cells (EPCs). In diabetic patients, both the number and function of EPCs are significantly reduced, leading to impaired vasculogenesis and angiogenesis. A decrease in the number of EPCs occurs due to oxidative stress and chronic inflammation in hyperglycemic conditions, impairing bone marrow’s capacity to release EPCs. Additionally, a chronic oxidative environment in DFUs restricts the release of pro-angiogenic markers like VEGF and stromal-derived factor-1α (SDF-1α), reducing EPC migration and homing to the wound. EPCs are essential for creating new blood vessels to deliver oxygen and nutrients to the injury site. Dysfunction and depletion of EPCs further compromise angiogenic responses, exacerbating impaired wound healing in DFUs [18–20].
The final stage of wound healing, known as the remodeling stage, begins approximately three weeks after the injury and can last up to a few months to a year (Figure 1). In this stage, the collagen that was present in the earlier stages of wound healing is switched from Type III collagen to Type I collagen. The collagen fibers are realigned along the lines of mechanical stress, which improves their tensile strength. However, the new tissue will only be about 80% of the original tensile strength [16]. The diabetic wounds get stuck in the inflammatory phase, and the healing response does not enter the proliferation phase (Figure 1).
The two most significant risk factors for the development of DFUs are DN and PAD. Patients affected by both conditions are at a substantially increased risk of developing DFUs. DN leads to a loss of protective sensation in the extremities, allowing minor injuries or repetitive trauma to go unnoticed and remain untreated for prolonged periods. Epidemiological data show that neuropathy is responsible for 16–66% of the cases of diabetic foot syndrome [2]. Even after ulcer healing, patients with neuropathy are prone to recurrence, which ultimately increases the risk of lower-limb amputation. PAD, which is characterized by the narrowing or obstruction of arteries in the periphery, most commonly affecting the lower extremities. These arterial blockages impair the flow of blood to the affected area, resulting in tissue ischemia. Inadequate perfusion compromises wound healing, reduces the ability to combat infection, and disrupts the maintenance of healthy skin and nerves. Consequently, patients with PAD face a significantly higher risk of infection and amputation, particularly when PAD coexists with DN [2]. These risk factors, if persistently present, not only increase the incidence and prevalence of DFUs but also contribute to non-healing chronic DFUs.
DFUs are primarily classified by their underlying cause into three main types: neuropathic (nerve damage, often on the sole), ischemic (poor circulation, often on toes/heels), and neuroischemic (combination of both). These chronic, slow-healing wounds require specialized care to prevent infection, which can lead to severe complications [1, 21].
Ischemic ulcers (approximately 15% [1]) arise when an area of the foot receives improper blood flow, often caused by stenosis of the arteries (PAD) without significant neuropathy. The impaired blood flow also leads to a decrease in nutrients in those areas, which causes damaged tissues to heal slowly. These are often found on the heels or tips of toes, frequently appearing painful, dry, and necrotic (blackened). Symptoms often include wounds on the feet that vary in color, which are typically raised around the edges, minimal/no bleeding, wounds that expose tendons, and dry/shiny/tight/hairless skin. Other observations include a person’s leg turning cold and pale when raising or turning red when dangling off a surface [2, 21].
Neuropathic foot ulcers (approximately 60% to over 85% of DFUs are considered neuropathic [22], but 35% are purely neuropathic [1]) occur because of impaired nerve function in the foot (peripheral neuropathy). Pain is lessened or absent in individuals with this type of ulcer, which means they don’t feel when their foot is being extensively damaged by repeatedly walking on certain areas of the foot exclusively. Since the weight is not being shifted or properly distributed, the damage worsens, and the skin begins to break down, eventually leading to the development of an ulcer. These commonly occur on the bottom of the feet (plantar surface) and are often surrounded by calluses [2, 21]. There are several clinical and molecular biomarkers that can predict DFU development in neuropathic patients. Key predictors include systemic inflammatory markers like neutrophil-lymphocyte ratio (NLR), elevated VEGF/intracellular adhesion molecule 1 (ICAM-1) for vascular dysfunction, and digital tools like plantar thermography to detect pre-ulcerative hotspots [23–26].
Neuroischemic ulcers (up to 50% [1]) are a combination of both neuropathic (loss of feeling) and ischemic (poor blood flow) issues. These are common in patients with long-standing diabetes. Diabetic under-the-skin/deep tissue (DUDT) ulcers: a distinct, sometimes traumatic, type that may not involve initial skin breakdown but indicates severe underlying damage. DUDT ulcers are serious, non-healing sores, often on the feet, that penetrate below the skin due to neuropathy, poor circulation, and pressure. They require immediate podiatric care to prevent severe infection, gangrene, or amputation. Key treatments include specialized debridement, off-loading pressure, infection control, and vascular assessment [2, 21].
DFUs arise from multiple interacting pathological processes, all driven primarily by chronic hyperglycemia. Understanding the molecular mechanism underlying DFU pathogenesis and non-healing pattern is important to understand because an in-depth understanding of various mediators will help identify the therapeutic targets regulating various molecular aspects, including inflammation, granulation tissue formation, ECM remodeling, and angiogenesis.
Poor management of DM results in chronic hyperglycemia, which disrupts multiple intracellular metabolic pathways, including the polyol and hexosamine pathways. Dysregulation of these pathways exacerbates oxidative stress, damages endothelial cells, delays wound repair, and contributes to the development of DN. Hyperglycemia also promotes the formation of AGEs, which accumulate on matrix proteins, disrupting cellular communications and delaying wound healing [4] (Figure 2).
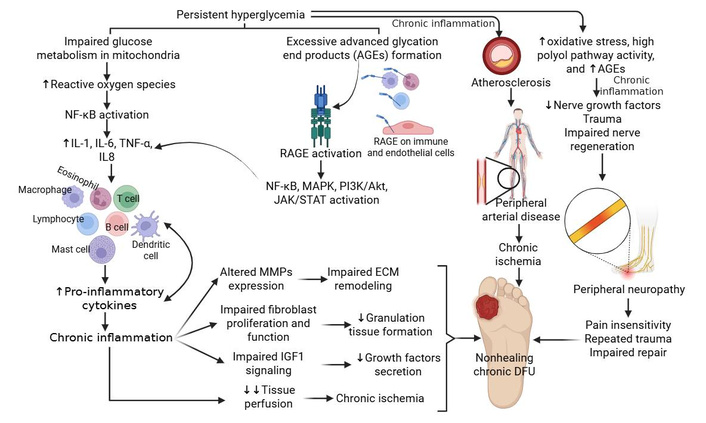
Molecular mechanisms underlying diabetic foot ulcer (DFU) pathogenesis and chronic non-healing. Chronic inflammation mediated by increased reactive oxygen species (ROS) and advanced glycation end products (AGEs) formation plays a critical role in the pathogenesis and chronicity of DFUs. It prevents healing by keeping the wound stuck in an active, dysfunctional inflammatory phase, often due to high blood sugar and ischemia. This persistent state features high levels of pro-inflammatory cytokines, excess proteases that degrade tissues, and ineffective macrophages that fail to clean the wound debris. Akt: protein kinase B; ECM: extracellular matrix; IGF: insulin-like growth factor; IL: interleukin; JAK/STAT: Janus kinase/signal transducers and activators of transcription; MAPK: mitogen-activated protein kinase; MMPs: matrix metalloproteinases; NF-kB: nuclear factor kappa B; PI3Ks: phosphoinositide 3-kinases; RAGEs: receptor of advanced glycation end products; TNF: tumor necrosis factor. Created in BioRender. Rai, V. (2026) https://BioRender.com/49arljc.
The roles of chemokine (C-X-C motif) ligand (CXCL8) (gene coding for IL-8), contributing to inflammation, angiogenesis, and phenotypic change of fibroblast [27], CD40-positive fibroblasts in inflammation and non-healing [28], fibroblast plasticity and heterogeneity [29], mediators regulating angiogenesis [30], proliferation, migration, and differentiation of fibroblasts, keratinocytes, and vascular smooth muscle cells [14] (Figure 2) during wound healing have been discussed in detail in our previous publications. Further, the role of macrophages, macrophage polarization, T cells, neutrophils, CXCL1, transforming growth factor (TGF)-β, IL-36γ, C-C chemokine receptor 4 (CCR4), TNF-α, IL-1, IL-6, IL-12, mitochondrial dysfunction, oxidative stress, decreased growth factors, matrix metalloproteinases (MMPs), immune response, phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) signaling pathway, Janus kinase/signal transducers and activators of transcription (JAK/STAT) signaling pathway, Wnt/β-catenin signaling pathway, Notch signaling pathway, and NF-κB signaling pathway in the pathogenesis of DFU non-healing have been discussed [12, 31–33] (Figure 2).
The AGE-RAGE (receptor of AGE) axis is an important contributor to DFU pathogenesis. This axis activates NF-κB, PI3K-Akt, and JAK/STAT signaling, promoting transcription factors and the release of inflammatory cytokines and increased expression of MMPs (Figure 2). All these factors contribute to impaired vascular function, immune regulation, and cellular regeneration, resulting in non-healing DFUs [34, 35]. While MMPs are essential for normal tissue remodeling, high levels of MMP-9, in particular, are strongly associated with poor DFU prognosis. Overexpression of MMP-9 in DFU tissues and wound fluid leads to excessive tissue degradation and prolonged inflammation. It acts as a primary detrimental factor in DFU recalcitrance. Unlike MMP-9, studies show that MMP-8 serves a beneficial role in DFU repair, aiding in collagen remodeling, whereas excessive MMP-9 destroys it. Overexpression of MMP-1, MMP-2, and MMP-13 is generally observed in non-healing wounds, contributing to the persistent inflammatory state. Thus, MMPs are critical gene targets in the treatment of DFUs, primarily because their overexpression in chronic wounds leads to excessive ECM degradation and inhibited healing [36–38] (Figure 2). It is important to note that the mediators mentioned above may be of therapeutic significance to promote healing.
DFUs typically begin as superficial skin lesions resulting from DN. DN has three key components that contribute to the formation of DFUs. 1) Sensory neuropathy involves the loss of protective sensation in the feet, including diminished pain, temperature, and pressure perception. This is the “primary link” to ulceration, where damage to nerves causes a “sock-like” loss of feeling in the feet. Patients cannot feel pressure, pain, or temperature changes, allowing minor injuries, like blisters, cuts, or burns from hot water, to go unnoticed and progress into serious, infected ulcers. 2) Motor neuropathy results from the denervation of intrinsic foot muscles, leading to muscle imbalance, foot deformities, and increased plantar pressure. These abnormal pressure points predispose the foot to repetitive trauma and subsequent infection. Abnormal pressure causes structural changes, such as claw toes or prominent metatarsal heads, which create new, abnormal pressure points. These areas are then subjected to repetitive, high-pressure trauma during walking, leading to skin breakdown and ulceration. 3) Autonomic neuropathy is characterized by impaired regulation of sweat and oil glands, resulting in dry, cracked skin that compromises the skin barrier and creates portals of entry for bacteria, leading to infections that often go unnoticed due to the concomitant sensory loss [3, 39]. In the process of DFU formation, the first stage is callus formation, which is driven by DN. Repeated mechanical stress causes subcutaneous bleeding beneath calluses, and progressive callus erosion ultimately leads to ulcer formation. This process is compounded by peripheral arterial disease. Reduced blood flow impairs oxygen and nutrient delivery, preventing normal wound healing and promoting tissue ischemia, which can progress to necrosis and gangrene [3]. The high glucose microenvironment also disrupts the normal healing process as it promotes bacterial growth and biofilm formation. Once bacteria invade the system, it can spread infection from the skin, subcutaneous tissue, fascia, and bone. As a result, osteomyelitis is a frequent complication of DFUs [12] (Figure 2). Our recent findings suggested altered expression of mediators of nerve regeneration, including activin A and synaptophysin, with hyperglycemia in a rat model of diabetes with ulcers [40]. These findings support the notion that impaired axonal regeneration in hyperglycemia after trauma is a contributing factor to the chronicity of diabetic ulcers.
PAD, a consequence of chronic inflammation mediated atherosclerosis, is highly prevalent in patients with DFUs (Figure 2), affecting up to 50–70% of cases and acting as a major cause of non-healing, infection, and amputation. PAD restricts blood flow to the feet, worsening the prognosis of ulcers and significantly increasing mortality risk, with 5-year survival rates sometimes lower than many cancers. PAD is an independent predictor of major limb loss in diabetic patients. Research indicates DFU healing rates are significantly lower in patients with PAD (69%) compared to those without (84%). Because diabetes often causes nerve damage (neuropathy), patients may not feel pain from ischemia, allowing the ulcer and underlying vascular disease to progress unnoticed [41, 42].
Biofilm formation in DFUs is a complex, molecularly driven process where bacteria shift from a planktonic to a sessile, matrix-enclosed state. Driven by hyperglycemia and chronic inflammation, key mechanisms include adhesion, extracellular polymeric substance (EPS) matrix production, quorum sensing, and genetic mutations that promote antibiotic resistance, impacting 70–80% of chronic wounds [43–45]. Specific mutations (e.g., in the mucA gene) in Pseudomonas aeruginosa drive excess alginate production, enhancing stability. Staphylococcus aureus often forms biofilms using ica-dependent pathways. The hypoxic (< 5 mmHg) and alkaline (pH 7.5–8.5) environment of a DFU stabilizes the biofilm, increasing its persistence. Finally, biofilms inhibit macrophage activation and block neutrophils, leading to persistent, non-healing inflammation. Pathogens like S. aureus (Gram-positive) and P. aeruginosa (Gram-negative) attach to wound tissue using surface adhesion proteins. The hyperglycemia in DFU stimulates this early adhesion. Targeting the initial adhesion phase (before matrix maturation) prevents bacteria from attaching via adhesins to wound surfaces [43–45]. These bacteria produce a protective matrix (polysaccharides, DNA, proteins) triggered by high glucose environments, which helps shield the biofilm from antibiotics and the host immune system. Using novel antiadhesion strategies may prevent the formation of a protective matrix and break the cycle of infection. The EPS matrix contributes to chronic infection, poor healing, and high amputation risk; targeting it may promote wound healing. Bacteria use chemical signaling [e.g., acyl-homoserine lactone (AHL) in Gram-negative, peptides in Gram-positive] to detect cell density. Once a quorum is reached, signaling pathways activate virulence genes and EPS production, allowing the biofilm to mature. Quorum sensing is a bacterial communication process that regulates gene expression based on cell-population density. Targeting quorum sensing aims to disrupt bacterial communication, reducing virulence and biofilm formation, particularly in P. aeruginosa and S. aureus [46, 47]. This suggests that biofilm formation plays a critical role in DFU non-healing; targeting biofilm will be of importance because it acts as a protective shield for bacteria, causing extreme resistance to antibiotics/antiseptics, perpetuating high inflammation, and directly causing tissue degradation, which can lead to infection progression, amputation, or death [48].
The above discussion suggests that chronic inflammation is a common denominator underlying the non-healing pathogenesis of non-healing DFUs. Since DFU is a multifactorial disease, it is not easy to specify any one pathway, but targeting inflammatory pathways to suppress chronic inflammation, which ultimately will promote angiogenesis, ECM remodeling, and granulation tissue formation, would be a potential target. Based on the above discussion, targeting chronic inflammation [IL-1, C-X-C motif chemokine receptor 2 (CXCR2), and TNF-α] may be an ideal candidate.
Current management and treatment for DFUs revolve around three strategies: debridement, offloading, and infection control. Debridement describes the process of removing necrotic/non-viable or hyperkeratotic tissue surgically. This process creates new wounds, which promote healing by accelerating the process of new tissue generation. It is considered a vital step in the healing process. Non-surgical methods involve less invasive treatments, including hydrogels, enzymatic debridement, and hydrotherapy. A risk of debridement is secondary infection, which delays the healing process further [49, 50].
Glycemic control is the process of controlling blood glucose levels to a degree that has been shown to delay/reduce the occurrence of DFU symptoms. HbA1c levels are increased when healing is decreased, and studies have shown that even a 1% drop is enough to reduce microvascular complications by as much as 37% [51–53]. Although using intense glycemic control can be beneficial, it may also lead to complications such as hypoglycemia. An additional issue with glycemic control is that an acute trial showed no correlation between meticulously controlling glucose levels and positive wound outcomes. Even so, hyperglycemia is a common reason for the recurrence of DFU, so maintaining glucose at adequate levels is essential [51–54]. Exercise is another effective method of treatment, because it decreases the risk of risk factors. All forms of exercise, when performed together, have shown the most promising results, although a clear relationship between exercise and decreased DFU rates has not been established [55].
Some novel treatments that do not directly involve gene editing but still show promising results are skin grafts and low-level laser therapy (LLLT). Skin grafting and advanced skin substitutes for DFUs work by altering the gene expression profile of the chronic wound environment, shifting it from a non-healing state to a healing phenotype. This involves reducing inflammation (specifically lowering IL-6, TNF-α, and MMP expression) and promoting angiogenesis and fibroblast proliferation by upregulating factors like VEGF, PDGFA, and TGF-β1 [56, 57]. Fish skin xenografts can provide an efficient and readily available [58]. One recent treatment that has been shown to improve DFU patient outcomes is using intact fish skin grafts. Atlantic cod fish is decellularized, lyophilized, and sent either intact or as mesh sheets. Patients who underwent this treatment underwent heavy debridement to remove all non-functioning tissue. The fish skin was applied while the wound was bleeding and then covered with a dressing. This was paired with consistent wound care (cleaning) along with off-loading. Patients who followed up after 16 weeks showed faster healing times when compared with the control group [59]. Skin grafting is a highly effective, advanced treatment for chronic DFUs that have not responded to conventional care, healing wounds 40–50% faster. It involves covering cleaned (debrided) ulcers with skin, either autologous or bioengineered, to promote rapid wound closure, prevent infection, and improve limb salvage rates [60–62].
Laser therapy, more specifically low-energy laser therapy, has been found to have stimulating effects on cells. LLLT changes gene expression by stimulating mitochondria to produce more ATP and reactive oxygen species (ROS), which act as signaling molecules to activate transcription factors like activator protein (AP)-1 and nuclear factor kappa B (NF-kB). This modulation typically upregulates pro-survival and tissue-repair genes while downregulating pro-inflammatory genes, accelerating healing, reducing inflammation, and boosting cell proliferation [63, 64]. LLLT promotes healing by stimulating cellular repair by increasing fibroblast production, which helps “knit” tissue back together, speeding up wound closure. By enhancing blood flow (microcirculation) around the injury, LLLT helps deliver nutrients and oxygen, aiding the removal of waste products. LLLT helps control chronic inflammation and can reduce infection rates, which is crucial for recalcitrant, non-healing DFUs. LLLT also helps optimize tissue regeneration rather than excessive scarring (Table 1). In cell studies with human skin, keratin, and fibroblast cells, exposure to low levels of radiation from the low-energy laser results in an increased production of TGF-β1 and MMP-2. When comparing cells irradiated at 1,064 nm and 632.8 nm, the cells exposed to the lower level of light, such as 830 nm (visible range), displayed increased rates of haptotaxis, migration, and ATP luminescence. These results indicate that low-energy radiation from a laser may increase the rate at which wounds close [65]. The studies in Table 1 evidence the beneficial effects of LLLT on wound healing; however, these studies have not discussed the change in gene expression after LLLT. Future studies are warranted to explore the change in genetic landscape and transcriptomic profiles during wound healing in DFU after LLLT.
Skin grafting and low-level laser therapy (LLLT) in the treatment of diabetic foot ulcers (DFUs).
| Study | Type of ulcer/patient population | Intervention | Results |
|---|---|---|---|
| Skin grafting | |||
| Saydam et al. [60] | A retrospective clinical study with 15 DFU patients. | AEG on DFUs. | AEG may be used effectively and safely in patients with DFU emergencies. |
| Shetty et al. [61] | An RCT with 52 patients (UTSG vs. conventional dressing). | UTSG was used on DFUs. | 84.61% of wounds managed with UTSG healed completely after 12-weeks compared to 53.84% of wounds managed with conventional dressing. |
| Tzeng et al. [62] | Eight diabetic patients with nine non-healing lower extremity ulcers. | Wound debridement, autologous PRP and thrombin, and thin split-thickness skin graft on the wound. | Complete wound healing in 2 to 3 weeks in 8 successful cases.No ulcer recurrence. |
| Laser treatment | |||
| Waluyo and Hidayat [66] | 55-year-old male patient with DM, diabetic neuropathy, and diabetic foot. | Normal saline, topical antibiotics, and LLLT, with a dose of 10 J/cm2, with a frequency of therapy of 3 times per week. | Wound tissue growth and no significant adverse effects after 12 weeks. |
| Santos et al. [67] | Thirteen RCTs with a total of 361 participants with DFUs. | Treatment with 632.8 to 685 nm, 50 mW/cm2, 3 to 6 J/cm2, and irradiation for 30 to 80 seconds, 3 times weekly. | Only 3 RCTs reported a reduction in the size of the ulcers.LLLT for a month is effective, safe, and of benefit for DFU treatment. |
| Huang et al. [68] | 13 RCTs and 413 patients with DFU. | LLLT was given to patients with DFUs.Various wavelengths in these 13 studies were 830 nm, 632.8 nm, 650 nm, 660 nm, 685 nm, 904 nm, and 400–800 nm. | LLLT significantly increased the complete healing rate, reduced the ulcer area, and shortened the mean healing time of patients with DFUs. |
| Chen et al. [69] | A meta-analysis of 11 studies involving 657 participants with grade I–II DFUs. | Various wavelengths in these studies were 660 nm, 630 and 810 nm, 400–800 nm, 635 nm, 890 nm, and 685 nm. | LLLT significantly increased the complete healing rate, the wound shrinkage area, and the wound bacterial clearance rate compared to the control group and reduced the infection rate. |
| Sani and Sani [70] | 20 patients with chronic DFUs. | LLLT and PRP therapy, along with standard wound care. | Two patients showed significant clinical improvement with resolution of necrotic tissue, reduction in wound size, and formation of healthy granulation tissue. |
AEG: autologous epidermal grafting; DM: diabetes mellitus; PRP: platelet-rich plasma; RCT: randomized controlled trial; UTSG: ultrathin skin graft.
Non-healing is one of the most significant complications in patients with DFUs. This may be due to factors including poor wound environment, preexisting health issues, and certain medications. Chronic inflammation, elevated protease activity of MMPs and serine proteases that negatively affect collagen synthesis and growth factors, cause non-healing in DFU [71]. The risk of non-healing increases the likelihood of amputation (with up to 30% of cases resulting in minor or major amputations). Certain risk factors, including gender, smoking, body mass index (BMI), hypertension, cardiovascular disease (CVD), kidney disease, white blood cell (WBC), hemoglobin, and albumin levels, all increase the likelihood of amputation [72].
Another common complication of patients with DFUs is the high recurrence rates. DFUs are long-lasting and slow-healing, increasing their sensitivity to infection. Even after healing completely, the area remains susceptible at 40% in the first year and up to 65% within 5 years. When recurrence occurs, the chances of complications arising increase exponentially [73].
DFUs also have complications of severe infections (cellulitis, abscess, osteomyelitis) and gangrene. Over 50% of ulcers become infected, leading to cellulitis (skin infection), deep tissue abscesses, and sepsis. Osteomyelitis (infection reaching the bone) occurs in up to 50% of cases. Tissue death (necrosis) due to lack of blood flow, often requiring urgent intervention, is gangrene of the foot, and severe, inflammatory structural deformity that can result from neuropathy leads to Charcot Foot. High 5-year mortality rates (up to 60% post-amputation) and increased risk of CVD are associated complications. Chronic pain, loss of mobility, and prolonged hospitalization reduce the quality of life [74]. This suggests that despite the existing treatment strategies, the existing issues of high cost, recurrence, amputation, and even death with DFU healing indicate the need for better modalities. Stem cell therapy [75] and gene therapy are emerging strategies for promoting DFU healing by delivering genetic material to enhance tissue regeneration, angiogenesis, and wound closure.
Gene-based medicine is the process of modifying the expression of genes or their properties for use in medical treatment. Genes that cause disease could be deactivated or could be activated to stop the disease. Also, ones that are not working could be replaced by healthy ones to treat specific diseases. It can also work by signaling to your body’s own immune system that a foreign intruder is present, allowing an immune response to occur [76]. Gene therapy utilizes vectors (often viruses) to deliver functional genes to correct faulty, missing, or mutated genes responsible for conditions like cancer, blood disorders, and inherited diseases. Viral methods (most common methos utilizing modified viruses like adeno-associated virus (AAV), Lentivirus that cannot cause disease but excel at entering cells) offer high efficiency and long-term expression but risk immunogenicity, while nonviral methods like lipid nanoparticles (LNPs) (increasingly favored for safety and scalability, often employing cationic lipids or polymers to encapsulate DNA/RNA), electroporation (create temporary pores in cell membranes), hydrodynamic delivery (rapid, high-volume injection to push DNA into tissues), and polymer-based vectors (like polyethyleneimine) to condense DNA for delivery offering flexibility in design offer greater safety, lower costs, and larger capacity [77–79].
Key methods include gene addition and gene editing (e.g., CRISPR), with treatments typically administered in vivo (directly into the body) or ex vivo (cells modified outside the body) [80]. By inserting functional genes into cells, gene therapy allows the body to produce necessary proteins, correct genetic defects, or enable the immune system to fight diseases more effectively. Gene replacement/addition (replacing a non-functional gene with a new, working copy), gene silencing/inactivation (turning off a mutated or malfunctioning gene), and gene editing (precisely repairing a broken gene) are different types of gene therapy. There are two main classes of gene-based medicine: in vivo and ex vivo. In vivo gene therapy describes the process in which humans are given agents directly to combat disease, while ex vivo describes the process in which human cells are genetically modified and injected back into the patient as a form of treatment [81].
Gene therapy provides one-time, long-term, or potentially curative treatments for previously incurable, severe diseases. However, gene therapy is also associated with potential risks and challenges, including immune reactions (the body may reject the vector (virus) delivering the gene due to an immune response), off-target effects (gene editing tools might unintentionally modify DNA in the wrong location), and infection (while rare, viral vectors could potentially cause infections) [82]. One way to attenuate vector rejection is by taking immunosuppressant drugs as well as delivering the cells outside of the patient’s body. Risks involved with gene-based therapy include an immune response where the vector is seen as foreign and attacked, leading to significant reactions such as organ failure. They may also affect cells that are healthy and do not require treatment. One additional risk includes the possibility of an error in the DNA, leading to cancer [82]. Another risk involved is that there is the possibility that the gene being introduced attaches itself to the wrong spot in the genome, which could vastly affect function and cause issues like cancer. Another problem with gene therapy is the costs involved, as genetic disorders are often rare, which means all of the resources put into a therapy may only be useful to a few patients. Although gene-based medicine is tailored for individuals, the costs involved are not viable, unlike mass-produced medicine such as vaccines [80, 83, 84].
The gene-based therapies that are currently being pursued include gene replacement, gene addition, gene slicing, gene editing, and gene activation/deactivation [81]. Gene-based therapies for DFU are emerging as advanced, targeted treatments designed to overcome chronic, non-healing wounds by inducing, inhibiting, or modifying specific gene expression. Gene-based medicine for DFUs utilizes engineered bacteria or DNA to deliver therapeutic genes encoding growth factors directly to the wound site. These approaches are designed to address key defects in diabetic wound healing, including reduced cellular activity and impaired growth factor signaling. By enhancing angiogenesis, stimulating tissue regeneration, and reducing inflammation, gene-based therapies promote blood vessel formation and tissue repair.
Angiogenic gene therapies for DFUs accelerate healing by delivering genetic material, typically for VEGF, FGF, or hepatocyte growth factor (HGF), to stimulate new blood vessel formation in chronic, oxygen-starved wounds. These therapies, often delivered via viral vectors or engineered scaffolds, improve tissue perfusion, reduce resting pain, and significantly enhance wound closure rates in preclinical and clinical studies [19]. Studies using VEGF mRNA delivery have demonstrated accelerated re-epithelialization and reduced wound area in diabetic mouse models. Two-stage gene therapy involving intramuscular injections of VEGF, HGF, and angiopoietin 1 (ANG1) plasmids has shown promise in treating critical limb ischemia, increasing perfusion, and decreasing pain [85]. Co-delivery of multiple growth factors (e.g., VEGF and PDGF-BB) has been shown to expand the therapeutic window and improve arteriogenesis [86]. Chronic inflammation plays a critical role in the chronicity of DFU, so it may be an attractive target for gene therapy. Anti-inflammatory gene modulation in DFU healing focuses on switching chronic, proinflammatory (M1) macrophage dominance to anti-inflammatory, reparative (M2) phenotypes, resolving persistent inflammation. Key strategies involve suppressing cytokines like TNF-α, IL-1β, and IL-6, while targeting genes that manage macrophage polarization, fibroblast function, and angiogenesis to reverse healing impairment [87, 88]. Currently, leading approaches to promote DFU healing include gene therapy using Lactococcus cremoris [AUP1602-C (AUP-16)], plasmid DNA expressing HGF (VM202), and nanoparticle-based delivery of miRNA (e.g., miRNA-146a) to target inflammation [89, 90].
AUP-16 is a gene-based therapeutic that employs genetically engineered L. cremoris, a non-pathogenic, probiotic bacterium, to locally deliver multiple human therapeutic proteins directly to the wound site in a targeted delivery. This engineered strain is designed to express and secrete basic FGF-2, IL-4, and macrophage colony-stimulating factor-1 (CSF1), all of which play critical roles in tissue repair and immune modulation [90] (Table 2). In the wound, AUP-16 aims to resolve chronic inflammation, stimulate neovascularization, and promote granulation tissue formation and re-epithelialization [90]. Upon application to the wound, the bacterium is recognized by the host immune system as a foreign stimulus, triggering a controlled acute inflammatory response. This immune activation serves as a “reset” converting the chronic inflammatory state characteristic of diabetic wounds into a productive acute inflammatory phase. The resulting recruitment of immune cells enhances wound surveillance and debris clearance, followed by the transition of the wound to the regenerative phase. Concurrently, FGF-2 and IL-4 stimulate fibroblast proliferation, ECM deposition, neovascularization, and keratinocyte activity, collectively supporting granulation tissue formation and epithelial repair [90]. The primary advantage of using AUP-16 is that it functions as a living bioreactor within the wound, employing a multi-target approach (IL-4, FGF-2, and MCSF-1) to rapidly heal complex, chronic, and neuro-ischemic ulcers with high efficacy. AUP-16 is safer and cost-effective than the current standard of care for DFUs.
Gene therapies for diabetic foot ulcers (DFUs).
| Study | Aim of the study/strategy | Patient population | Outcome |
|---|---|---|---|
| Schindler et al. [90]Phase I, open-label, dose-finding study | Four cohorts of patients received AUP1602-C as a single dose of 2.5 × 105 colony-forming unit (CFU)/cm2 ulcer size or as repeated doses between 2.5 × 106 and 2.5 × 108 CFU/cm2 administered 3 times per week for 6 weeks. | 16 patients aged 53-80 years. | AUP1602-C is safe and well-tolerated and showed dose-dependent efficacy in patients with DFU.A dose of 2.5 × 108 CFU/cm2 showed complete healing in 83% of patients.No recurrence in follow-up. |
| Perin et al. [89]Phase 3 study | 0.25 mg per 0.5 mL per injection via 16 injections (4 mg total per visit) of VM202, a plasmid DNA expressing two isoforms of human hepatocyte growth factor, into the ipsilateral calf of the affected foot on days 0, 14, 28, and 42. | 44 subjects with neuroischemic DFUs. | Wound closure in the VM202 group occurred from 3 to 6 months, but with no statistical significance.Intramuscular injections of VM202 plasmid DNA to calf muscle may have promise in promoting healing in chronic neuroischemic DFUs. |
| Kessler et al. [91]Randomized, placebo-controlled phase III study | To check the efficacy of VM202 in subjects with painful diabetic peripheral neuropathy. | Part I: 9 months with 500 subjects. Part II: 101 subjects with a noninterventional extension to 12 months. | VM202 failed to meet its efficacy endpoints in part I but showed significant and clinically meaningful pain reduction versus placebo in part II.VM202 may attenuate disease progression. |
| Dewberry et al. [92]Pre-clinical study | Cerium oxide nanoparticle conjugation to microRNA-146a. | Diabetic mice model. | Cerium oxide nanoparticle -microRNA-146a promotes wound healing. |
VM202 (ENGENSIS) is a genetically engineered plasmid DNA that has been modified to express HGF728 and HGF723. HGF is a naturally occurring growth factor that has been proven to promote angiogenesis and has also been shown to support nerve damage repair, cell survival, and decreased inflammation. VM202 is typically given in the calf muscle. DFUs are known to have issues like decreased blood flow, poor cell turnover, and nerve damage [89] (Table 2). VM202 was also found to be effective in reducing pain in painful diabetic peripheral neuropathy [91], a risk factor for DFUs. In animal models, intramuscular injection of HGF has shown increased angiogenesis in PAD [93], a contributor to DFU healing (Figure 3). VM202 has the advantage of increased nerve regeneration, an excellent safety profile, long-lasting therapeutic benefits (decreasing the cost of conventional DFU treatment), and its effectiveness in neuroischemic ulcers.
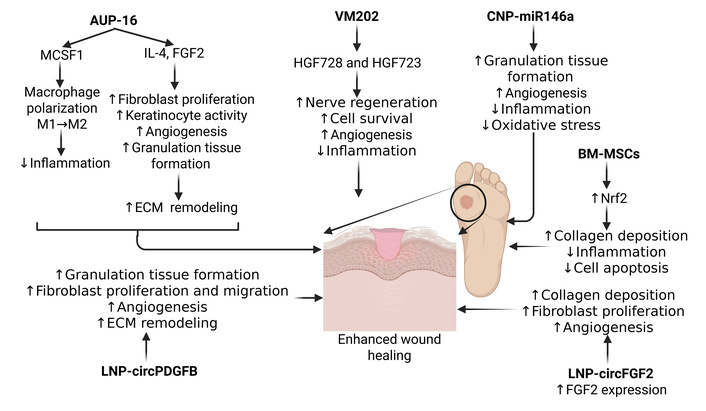
Gene therapies for diabetic foot ulcer healing. LNP: lipid nanoparticle; Nrf2: nuclear factor erythroid 2-related factor 2. Created in BioRender. Rai, V. (2026) https://BioRender.com/nrk14fd.
While plasmid DNA (VM202) is often used due to its simplicity and safety, bacterial vectors (AUP-16) offer advantages in terms of delivery efficiency, immunogenicity, and target specificity. Naked plasmid DNA, like VM202, often suffers from low efficiency in entering human cells. Bacterial vectors (e.g., Salmonella, Listeria, Lactococcus) are naturally proficient at invading mammalian cells and delivering their genetic cargo directly to the cytoplasm. Bacterial vectors provide an inherent adjuvant effect, stimulating both cellular (CD8+ T cell) and humoral (antibody) immune responses more effectively than naked plasmid DNA. Bacterial vectors can be administered via non-invasive routes, allowing for targeted mucosal immunity, whereas plasmid DNA usually requires direct injection [94, 95]. Along with VM202, two-stage gene therapy combining angiogenesis-stimulating plasmids (VEGF, HGF, ANG1), nerve growth factor plasmid (EGF), and nanoparticle-delivered plasmid are other plasmid-based therapies for DFU, as summarized in Table 3.
Summary of the plasmid based gene therapies to promote diabetic foot ulcer (DFU) healing.
| Study | Aim of the study/phase | Subjects/intervention | Outcome |
|---|---|---|---|
| Perin et al. [89] | A phase 3, double-blind, randomised, placebo-controlled, multicenter 7-month study for the efficacy and safety of VM202. | 44 patients with DFU.VM202 was injected via 16 injections (4 mg total per visit) into the ipsilateral calf of the affected foot on days 0, 14, 28, and 42. | 23 patients with neuroischemic ulcers: significantly increased complete ulcer closure at months 3, 4, and 5 (P = 0.0391, 0.0391, and 0.0361).Increase in ABI in the treatment group.Phase 3 was discontinued due to slow patient recruitment, mixed efficacy results in earlier studies, and a need for further validation to prove it outperforms existing standard care. |
| Barć et al. [85] | A clinical study to evaluate the safety and efficacy.A phase of therapeutic trial. | 12 patients with gene therapy (treatment group).12 patients without gene therapy.First injection of pIRES/VEGF165/HGF followed by ANG1 plasmid injection after one month in treatment group. | Significantly improved healing in the treatment group associated with increased ABI, PtcO2 and decreased resting pain.Not in clinics yet because of high manufacturing costs, regulatory complexities, safety concerns regarding gene delivery, and the need for more robust, large-scale clinical trial data. |
| Chung et al. [96] | In-vitro study to evaluate the effects of EGF. | Multiple cell linesMultiple human EGF isoforms were transfected. | EGF828 including a membrane-anchoring domain was released as mature EGF protein.EGF plasmid may be effective as non-viral gene therapy.Not in clinics due to the lack of approved, large-scale clinical trials proving safety and efficacy, high development costs, and the technical challenges of delivering nucleic acids to chronic wounds. |
| Ko et al. [97] | In-vivo study to compare EGF and VEGF effects on wound healing in diabetic C57BL/6J mice. | FRM-EGF828 or minicircle VEGF165 were prepared using p2øC31 plasmid and a mixture of minicircle-VEGF165 (20 µg) and pβ-EGF828 (20 µg) with microbubble solution (100 µg) was injected subcutaneously. | Accelerated wound therapy with gene therapy.VEGF was better than EGF for angiogenesis and increased blood circulation.Wound healing rates and histological findings were more accelerated in the EGF gene therapy compared to VEGF.Not in clinics due lack of lack of approved, large-scale clinical trials proving safety and efficacy. |
ABI: ankle-brachial index; ANG1: angiopoietin 1; HGF: hepatocyte growth factor; PtcO2: transcutaneous oxygen pressure measurement; VEGF: vascular endothelial growth factor.
Another form of gene therapy is to target miRNAs (nanoparticle-delivered miRNA). Targeted therapies that utilize nanotechnology to deliver miRNA-146a to reduce excessive inflammation, a key barrier to healing in diabetic wounds [92]. Dewberry et al. [92] reported that intradermal injection of conjugated nanoparticle-miR146a promotes diabetic wound healing by increasing wound collagen and angiogenesis, while lowering inflammation and oxidative stress (Table 2, Figure 3). An emerging, non-viral approach using LNP and circular RNA (circRNA) to deliver long-term FGF-2 expression, accelerating wound healing by enhancing angiogenesis and collagen deposition, is another strategy of gene therapy. By protecting the RNA from degradation, this platform enhances angiogenesis, fibroblast proliferation, and collagen deposition, offering a superior “single-dose, long-acting” alternative to conventional protein therapies for diabetic, chronic wounds. The LNP-circFGF-2 system delivers circRNA into cells, which then translates into functional FGF-2 protein. Due to the circular structure of RNA, it is highly resistant to exonucleases, resulting in prolonged expression compared to linear mRNA. The sustained production of FGF-2 promotes angiogenesis (new blood vessel formation), increases fibroblast migration/proliferation, and encourages collagen remodeling [98] (Figure 3). Unlike directly applied protein, which degrades quickly in the harsh, high-protease environment of chronic wounds, the LNP-circRNA approach enables sustained, robust protein production from within the host cells. Studies have shown that LNP-circFGF-2 effectively accelerates wound healing in diabetic mice, addressing the need for more effective, long-acting treatments for DFUs. The use of ionizable lipids for formulation allows efficient delivery while maintaining low immunogenicity, making it a promising non-viral gene therapy [98]. Another study documented that LNP-encapsulated PDGFB circRNA (LNP-circPDGFB) formulation delivered to the wound site promotes proliferation, migration, and ECM deposition of fibroblasts, and angiogenesis, enhancing diabetic wound healing [99] (Figure 3). Nanoparticle-delivered miRNA excels in high-efficiency, targeted, transient regulation of multiple gene targets simultaneously. While comparing the cost of bacterial vectors vs. plasmid DNA vs. nanoparticle-delivered miRNA, generally, bacterial vectors are the least expensive to produce, followed by plasmid DNA, with nanoparticle-delivered miRNA/siRNA being the most costly due to complex synthesis and formulation processes [99].
Stem cell-gene combination may be another strategy of gene therapy to promote wound healing in DFUs. Research indicates that strengthening bone marrow-derived mesenchymal stem cells (BM-MSCs) by modulating genes like nuclear factor erythroid 2-related factor 2 (Nrf2) (which increases antioxidant capacity) can enhance DFU healing by reducing oxidative stress-induced cell apoptosis. Nrf2 is a key transcription factor that boosts the antioxidant defense system [e.g., superoxide dismutase (SOD) and heme oxygenase-1 (HO-1)] in MSCs. Overexpression of Nrf2 protects BM-MSCs against high-glucose, hypoxic, and oxidative stress conditions, significantly reducing the apoptosis (cell death) rate of transplanted cells in the harsh DFU microenvironment (Figure 3). Nrf2-modified MSCs (and their derived exosomes) enhance angiogenesis, promote fibroblast migration, and accelerate tissue regeneration. Combining MSC therapy with Nrf2 activators (e.g., pharmacological agents) has been shown to result in superior wound closure, increased collagen deposition, and reduced inflammation compared to using MSCs alone [75, 100–102]. Stem cell-gene combination increases the effectiveness of regenerative medicine and improves the efficiency of genetic therapies, though it faces significant challenges, including technical complexity and cost.
As discussed above, gene therapy for DFU offers significant potential to accelerate wound healing and reduce amputations, particularly for neuroischemic ulcers that fail to respond to standard care. By delivering therapeutic genes to promote angiogenesis and tissue regeneration, this approach targets the underlying causes of chronic wounds, with novel therapies like VM202 showing promising results in phase 3 trials. The following are the key benefits of gene therapy in DFU. i) Accelerated wound closure: Gene therapy, such as VM202, has shown significant potential in promoting the closure of chronic DFUs, especially in patients with neuroischemic conditions. ii) Key mechanisms involve delivering genes that stimulate the growth of new blood vessels, addressing the poor local blood circulation that prevents wound healing. iii) Reduced amputation rates: by improving healing, this therapy offers a potential alternative to surgical intervention, aiming to reduce the high rate of amputations associated with chronic DFUs. iv) Targeted therapeutic delivery: gene therapy allows for the direct application of growth factors and other necessary proteins to the wound site, often using plasmid DNA or other vectors to enhance healing. v) Improved clinical outcomes: Studies have indicated a positive trend in wound reduction, with some patients achieving complete, sustained closure of ulcers that were previously resistant to treatment [75, 89, 91, 103].
Gene therapy addresses significant unmet needs in DFU treatment, such as chronic non-healing, infection, and amputation risks, by enabling local, sustained production of therapeutic proteins directly at the wound site [104]. Innovative approaches like AUP-16 use genetically engineered bacteria to deliver multiple growth factors, surpassing the limitations of traditional, short-lived protein therapies. As mentioned above, emerging therapies, such as AUP16, combine factors like FGF-2, IL-4, and CSF1 to address multiple aspects of wound healing. Another advantage of gene therapy is multi-target approach. A multi-target gene therapy approach for DFU aims to simultaneously address chronic inflammation, poor vascularization, and impaired healing. Leading candidates like AUP16 utilize engineered bacteria to deliver multiple therapeutic proteins (e.g., FGF-2, IL-4, CSF1) directly to the wound, demonstrating up to 83% healing rates [90]. Promising efficacy and safety are other advantages of gene therapy.
Multi-omics integrates genomics, transcriptomics, proteomics, and epigenomics to provide a holistic view of disease mechanisms, enabling precise target identification (e.g., receptors, enzymes, signaling pathways by revealing genetic, epigenetic, and metabolic alterations) and patient identification and stratification (by identifying predictive biomarkers to select patients who are more likely to respond to specific gene therapies) for gene therapy. By applying single-cell techniques, this approach optimizes therapy development, improves safety by detecting off-target effects (by understanding the molecular heterogeneity between patients), and enhances understanding of therapeutic efficacy. Multi-omics has various applications. Multi-omics identifies specific molecular subtypes of diseases, allowing for the selection of patient populations most likely to respond to gene therapies (precise patient stratification). Single-cell multi-omics, such as the Tapestry platform by Mission Bio, enables researchers to analyze thousands of cells simultaneously to detect unwanted off-target effects and confirm the quality of the final therapeutic product (safety and efficacy monitoring). By combining data layers, such as linking genetic variants (genomics) to gene expression changes (transcriptomics) and protein levels (proteomics), researchers can identify the most effective, or “causal” targets for intervention (target identification). Multi-omics analysis helps to uncover how therapies work at a molecular level, allowing for the optimization of drug combinations and the mitigation of resistance [105–107]. In the context of gene therapy, once a genetic target has been identified using multi-omics, the predicted target’s or gene perturbation’s effects may be validated using CRISPR-based knockdowns or overexpressions on disease pathways. Multi-omics accelerates the identification of druggable targets through automated, high-throughput computational tools, and this will help in developing targeted therapies. Technologies like CRISPR and functional vectors can be guided by multi-omics to improve the targeting and efficacy of gene editing, such as with reversible epigenetic editors [108, 109].
AI, by analyzing diverse data sources (genomic, transcriptomic, and proteomic), utilizes machine learning and bioinformatics to identify genetic targets (target identification by identifying disease-driving genes and predicting molecular interactions) in gene therapy, optimizing personalized treatments to promote healing in chronic, non-healing wounds. AI enhances target validation through 3D protein structure prediction (e.g., AlphaFold—an AI program for predictions of protein structure). These tools accelerate gene therapy development by optimizing CRISPR-based gene editing. This reduces off-target effects and models cell behavior to identify novel, druggable, and personalized gene targets for diseases. Artificial learning (AL) may be useful in various ways. Bioinformatics and machine learning [e.g., LASSO (least absolute shrinkage and selection operator)—a regression analysis method used for variable selection and regularization to enhance prediction accuracy, SVM-RFE (support vector machine recursive feature elimination)—a backward feature selection algorithm used in bioinformatics to identify relevant biomarkers when the number of features exceeds the number of observations, mitigating overfitting] analyze DFU-related gene expression data (GEO database) to identify key genes and signaling pathways for therapeutic intervention (target identification), such as targeting CXCR4 for tissue repair. AI ranks potential targets based on their likely therapeutic impact, helping researchers focus on the most effective genes to edit or regulate. This helps in predicting protein structures, allowing researchers to understand how pathogenic variants disrupt normal function and identify which components are suitable for gene modulation. AI also helps in integrating data from DFU images to improve management and treatment strategies [110–116]. Integrating patient-level genetic information, such as SNP data from the NOS1AP gene, with AI models predicts whether a DFU will heal (predictive prognosis). AI analyzes single-cell transcriptomics to understand stromal and immune cell behavior, identifying fibroblasts with stem-like, pro-inflammatory properties essential for healing (molecular imaging and cellular analysis). Machine learning optimizes gene-editing tools (e.g., TALENs, CRISPR) for precise, targeted genetic modifications (gene editing optimization to reduce off-target effects). Generative AI generates synthetic images for training and improved diagnostic segmentation, while smartphone apps use computer vision to monitor wound healing in real-time (supportive AI in management). Using graph neural networks, AI can map disease-related cellular pathways to determine which genes or protein interactions could be targeted to shift diseased cells back to a healthy state [110–116].
In regard to DFU healing, AI-powered models have transformed care by enhancing classification accuracy up to 95–97%, enabling early, automated detection via smartphone imaging, and predicting healing outcomes. These technologies analyze wounds to reduce clinician workload, standardize care, and provide real-time monitoring, crucial for preventing infection and amputation. AI analyzes wound photos to delineate boundaries and classify DFU severity using systems like Wagner’s grading with high precision. AI-powered smartphone applications allow for remote monitoring, enabling consistent, cost-effective follow-up without frequent clinic visits. Not only in classification and evaluating photos, AI evaluates infrared thermograms, electronic health record data, and biomechanical data for a holistic patient view [113, 114, 117, 118]. The role of AI and multi-omics and its beneficial results are supported by the fact that AI and multi-omics were involved in the development and design optimization of AUP16 [119]; however, there is no direct evidence that multi-omics or AI was specifically involved in the initial design or development of VM202. However, the development of AI and multi-omics methods may be useful in identifying potential targets for gene therapy, and these techniques must be incorporated in future studies.
Emerging therapeutic targets for DFUs focus on addressing chronic inflammation, impaired angiogenesis, and excessive fibrosis to promote healing. Key emerging targets include the prostaglandin I2 receptor (PGI2; regulates angiogenesis), CXCR4 (regulates stem cell mobilization and impaired angiogenesis), SCUBE1 (regulates immune regulation and inflammatory response), and RNF103-CHMP3 (regulates extracellular interactions and cellular communication). Research is also advancing in utilizing hypoxia inducible factor 1α (HIF-1α) for hypoxia reversal, Hippo-associated factors, the yes-associated protein (Hippo-YAP) signaling for macrophage polarization, and stem cell-derived exosomes for regeneration (Table 4, Figure 4). It should be noted that the studies listed in Table 4 are either ex vivo, pre-clinical, or network analysis but have not progressed to clinical trials.
Emerging genes as therapeutic targets for gene therapy in diabetic foot ulcers (DFUs).
| Target | Model | Strategy/approach | Outcome |
|---|---|---|---|
| PGI1 (integron class 1 gene)[120] | 50 specimens from DFU patients | Sample analysis revealed significantly increased integron class 1 gene in Proteus mirabilis isolated from diabetic foot infections. | The analysis of 50 DFU specimens showed a high prevalence of class 1 integrons among patients with P. mirabilis isolates.The PGI1 pathway may have therapeutic significance. |
| CXCR4 [110]CXCR4 [121] | GEO database analysis (DFU patient data) | The study identified differentially expressed genes and performed enrichment analysis to reveal the related biological pathways. | GEO database analysis revealed an important role of CXCR4 and its related pathways in the pathogenesis of DFUs [110]. CXCR4 also emerges as a potential target for intervention, along with PGI2, CSF3R, and CSF2RA, in clinical trials analysis of DFU [121].CXCR4 may be a potential target in DFU management. |
| Hippo-YAP signaling pathway [122] | Single-cell RNA sequencing of tissues from a diabetic mouse model | Mouse models were used to induce ulcers, and tissues were characterized histologically. Sequencing analysis was conducted to investigate the role of the TFAP2A-LIFR-Hippo-YAP signaling axis in regulating macrophage M2 polarization and its critical function in DFU wound healing. | Bulk and single-cell RNA sequencing of diabetic wounds revealed the critical role of macrophage M2 polarization mediated by TFAP2A, a central regulatory gene in macrophage function, in DFU healing.TFAP2A-LIFR-Hippo-YAP signaling axis accelerates DFU wound healing through the induction of macrophage M2 polarization.The immunoregulatory role of TFAP2A in DFU makes it a promising therapeutic target. |
| YAP-TAZ signaling [123] | Tissues from the ear wounds of diabetic mice | Gene and protein expression analyses on control and diabetic tissues.The aim was to evaluate the cellular regulation of Hippo and related pathway targets after plasma treatment. | The analysis of ear wounds in diabetic mice treated with cold plasma showed that cold plasma accelerated healing by increasing re-epithelialization and modifying extracellular matrix components involving YAP-TAZ signaling.YAP-TAZ signaling and activated TGF-β-regulated cellular regulation after gas plasma treatment have therapeutic potential, and YAP-TAZ is an attractive target. |
| SCUBE1 and RNF103-CHMP3 [124] | GEO database analysis of 15 DFU human samples | Differential gene analysis.Enrichment analysis.Protein-protein interaction network analysis. | SCUBE1 and RNF103-CHMP3, identified as key genes significantly associated with DFU, may be potential biomarkers and therapeutic targets. |
CXCR4: C-X-C motif chemokine receptor 4; GEO: Gene Ontology; Hippo-YAP: Hippo-associated factors, the yes-associated protein; LIFR: leukemia inhibitory factor receptor; TFAP2A: transcription factor AP-2 alpha; TGF: transforming growth factor; YAP-TAZ: yes-associated protein-transcriptional co-activator with PDZ binding motif.
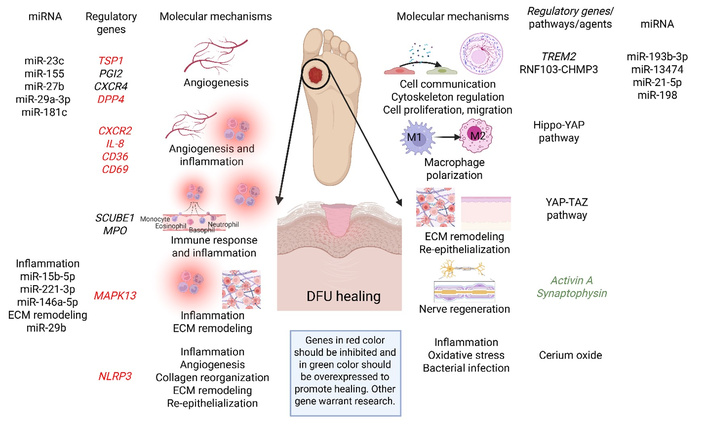
Emerging therapeutic targets to promote wound healing in diabetic foot ulcers (DFUs) and the molecular mechanisms regulated by these targets. Genes, pathways, and agents discussed in Targeting differentially expressed genes and Targeting miRNAs. CD: cluster of differentiation; CXCR4: C-X-C motif chemokine receptor 4; DPP4: dipeptidyl peptidase 4; ECM: extracellular matrix; Hippo-YAP: Hippo-associated factors, the yes-associated protein; IL: interleukin; MAPK13: mitogen-activated protein kinase 13; miR: microRNA; MPO: myeloperoxidase; NLRP3: NLR family pyrin domain containing 3; PGI2: prostaglandin I2 receptor; TREM2: triggering receptor expressed on myeloid cells 2; TSP1: thrombospondin 1; YAP-TAZ: yes-associated protein-transcriptional co-activator with PDZ binding motif. The genes in red should be targeted for inhibition/attenuation while in green should be promoted to promote wound healing. Created in BioRender. Rai, V. (2026) https://BioRender.com/25x5xvd.
As mentioned above, these emerging targets are involved in various molecular mechanisms (angiogenesis, inflammation, ECM remodeling, and cell proliferation and movement), which are involved in DFU healing, and targeting them alone or in combination may have therapeutic effects. Targeting multiple molecules in DFU healing is a therapeutic approach designed to address the complex, chronic, and non-healing nature of these wounds. This notion is supported by the findings of the synergistic actions of targeting multiple signaling pathways in promoting wound healing in DFU [125]. The notion of targeting these mediators is supported by their interaction (Figure 5A) and their involvement in various molecular mechanisms involved in wound healing (Figure 5B).
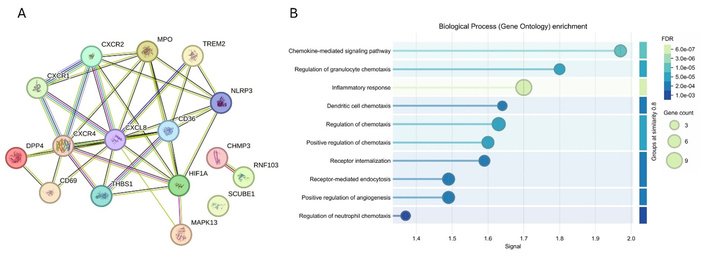
Protein-protein interaction between different mediators (panel A; as discussed in this section) and their involvement in various molecular mechanisms involved in wound healing (panel B). Networking was done using STRING 5 and 6 to show interactions between different mediators and molecular mechanisms involved in diabetic foot ulcer (DFU) healing to address this concern. https://string-db.org/.
In addition to the targets discussed in Table 4, studies have documented other potential therapeutic targets. For example, Yu et al. [126] with proteomic analysis of 54 DFU patients and age- and sex-matched healthy controls, reported differentially expressed proteins LRG1, CD5L, CRP, IGHA1, and LBP in DFU (regulating ECM remodeling, immune response, and complement activation) (Figure 4). These findings suggested that targeting these proteins may have therapeutic significance in promoting DFU healing [126]. Targeting dipeptidyl peptidase 4 (DPP4) has been suggested to promote DFU healing (regulating angiogenesis) using a meta-analysis of 4 randomized controlled trials (Figure 4). The results revealed significantly increased complete ulcer healing [risk ratio (RR) = 1.63, 95% CI 1.30–2.06, P < 0.0001] and shortened healing time [mean difference (MD) = –10.72 days, 95% CI –14.61 to –6.84, P < 0.00001] with DPP4 inhibitors [127, 128]. The study also revealed a decrease in the size of the ulcer (MD = –2.36 cm2, 95% CI –2.95 to –1.77, P < 0.00001). Tang et al. [129], using a rat model, reported that nicotinamide mononucleotide-modified CeO2 hydrogels promote wound healing (26.6% by day 4, 98.7% by day 10) in DFU (regulating inflammation) (Figure 4). CeO2 hydrogels also significantly decreased bacterial counts compared to control groups. These findings highlight the role of CeO2 in DFU healing by attenuating inflammation and oxidative stress by decreasing secretion of IL-1β and IL-18 [129]. Suppression of NLR family pyrin domain containing 3 (NLRP3) inflammasome-mediated inflammation in a rat model has been reported to promote wound healing in DFU. The finding revealed that NLRP3 inflammasome inhibitor Tan IIA significantly accelerated wound closure, enhanced re-epithelialization, improved collagen organization, and increased angiogenesis in diabetic rats [130] (Figure 4). Recent studies by our group using a rat model of diabetes with diabetic ulcers reported the increased expression of MAPK13, TSP1, and CXCR2 [131], CD34, and CD69 [132] (modulating angiogenesis, inflammation, and ECM remodeling), and decreased expression of activin A and synaptophysin [40] (modulating nerve regeneration) in DFU tissues of rats (Figure 4). Li et al. [133], through integrated multi-omics analysis, reported myeloperoxidase (MPO) (regulating immune response and inflammation) and triggering receptor expressed on myeloid cells 2 (TREM2) (cell cycle and cytoskeletal regulation) as secretory biomarkers and potential therapeutic targets (Figure 4). These results, though only a few among various published, suggest that differentially expressed genes and proteins may be potential targets for gene therapy; however, these targets warrant further research.
miRNAs, regulators of gene expression, are emerging as critical, druggable targets for DFUs by regulating pathways involved in inflammation, angiogenesis, and wound closure. Key miRNAs, including miR-193b-3p, miR-23c, and miR-155, are often dysregulated, impairing healing; targeting these with inhibitors [anti-miRNA oligonucleotides (AMOs)] or mimics can promote tissue repair [134, 135] (Table 5, Figure 4). The role of miRNA in promoting DFU healing is supported by the findings of the study of Dewberry et al. [92] using LNPs with miRNA-146a in reducing excessive inflammation and promoting wound healing in diabetic wounds (Figure 4). miR-181c plays a critical role in impaired angiogenesis in diabetes and is a therapeutic target for diabetic vascular complications [136]. Impaired angiogenesis is a causal factor for DFU, and thus miR-181c may have implications and therapeutic targets in DFU. miR-198 is a miRNA that acts as an anti-migratory factor, with elevated levels in non-healing DFUs, preventing proper skin repair by inhibiting keratinocyte migration. Researchers from A*STAR and Celligenics are developing AMOs to inhibit miR-198 (regulating ECM remodeling), aiming to accelerate healing and reduce amputations in DFU patients [137] (Figure 4).
MicroRNA as a therapeutic target to promote wound healing in diabetic foot ulcers (DFUs).
| miRNA | Model | Approach | Results |
|---|---|---|---|
| Cell proliferation, migration, function, and cytoskeleton | |||
| miR-193b-3p [138] | Genomic analysis of DFU tissue samples (n = 15 per group), in vitro keratinocyte (HaCaT) migration assays, organotypic 3D human skin wound models, and in vivo mouse wound healing model (n = 3 per group) | miRNA expression profiling, functional gain and loss of function assays, migration analyses, RhoA activity assessment, bioinformatic target prediction, and luciferase reporter validation to determine the mechanistic role of miR193b-3p and its regulation of KRAS | miR-193b-3p was found to be upregulated in DFUs, where it inhibited keratinocyte migration and reepithelialization while directly targeting KRAS and other oncogenic pathways. Contributing to impaired wound healing and reduced malignant transformation. |
| miR-13474 [139] | Exosomes secreted by HucMSCsDFU SD rats (n = 6 per group) received HucMSC exosomesRNA sequencing analysis of tissues | Blocking miR-13474 in HucMSC-derived exosomes | miR-13474 was significantly differentially expressed between the wound area and the wound edge area.miR-13474 targets the CPEB2/TWIST1 axis to improve the impaired function of skin cells |
| miR-21-5p [140] | In vitro studies | EVs were isolated from HucMSCs | miR-21-5p were overexpression in EVs.miR-21-5p inhibits KLF6 and promotes proliferation and migration of HSFs. |
| Inflammation | |||
| miR-15b-5p [141] | Human (14 DFU and 12 control) and swine DFU model (n = 2 in each group) | Transcriptome analyses of DFU tissue | S. aureus-triggered miR-15b-5p induction suppresses inflammation and DNA repair-related genes IKBKB and WEE1. |
| miR-221-3p [142] | Mouse DFU model (n = 6–10 in each group) and human keratinocytes (HaCaT) exposed to high levels of glucose | miR-211-3p levels increased, and the gene was knocked out. Measured inflammation and wound healing | miR-221-3p reduces inflammation and improves wound healing by suppressing DYRK1A, which decreases STAT3 activation and decreases inflammatory cytokine production. |
| ECM remodeling | |||
| miR-29b [143] | 32 patients with DFU (18 males and 14 females, aged 57.03 ± 8.23 years)Cerium dioxide nanoparticles-plasmid complexesDiabetic mouse model (n = 6 per group) | Microarray-based analysis in human samples followed by in-vtro and in-vivo analysis | DFU ulcerative edge in mice showed high expression of lncRNA H19 and FBN1 and low expression of miR-29b.lncRNA H19 and miR-29b promote wound healing involving FBN1. |
| Angiogenesis | |||
| miR-23c [144] | 40 subjects with normal glucose tolerance (n = 10), T2DM patients free from other complications, T2DM subjects with uninfected DFU, and T2DM subjects with infected DFU (n = 10 each) | hsa-miR-23a, hsa-miR-23b, hsa-miR-23c, and angiogenic factors such as SDF-1α, HIF-1α, VEGF, and eNOS were investigated in peripheral blood mononuclear cells and tissue biopsy samples using qPCR | SDF-1α has a significant inverse association with miR-23c.miR-23c functions as a new regulator to inhibit angiogenesis by targeting SDF-1α.. |
| miR-155 [145] | Human DFU patients’ wound tissuecontrol group (n = 36) and the PRF group (n = 24) | PRF treatment; miR-155 expression analysis | PRF treatment suppressed miR-155, increased HIF-1α /VEGF, and vascular density, as well as accelerated wound healing. |
| miR-27b [146] | Human chronic DFU (n = 10) tissue vs. acute DFU tissue (n = 12) and hyperglycemic endothelial cells | miR-27b and angiogenic marker expression analysis; knockdown of miR-27b and Nrf2 activation studies; miR-27b and Nrf2 pathway expression analysis | miR-27b levels were lower in chronic DFU and showed a correlation with Nrf2. Reducing miR-27b impaired Nrf2 signaling and angiogenesis. Activating Nrf2 restored miR-27b levels and gene expression. |
| miR-146a-5p (inflammation)and miR-29a-3p (angiogenesis) [147] | HaCaT cells | HaCaT cells were transfected with miR-146a-5p or miR-29a-3p inhibitors individually or in combination, following stimulation with TNF-α | miR-146a-5p and miR-29a-3p inhibitor supplementation to diabetic wounds reduced the wound size on days 8 and 9 but did not lead to complete healing by day 10. |
DFU: diabetic foot ulcer; ECM: extracellular matrix; eNOS: endothelial nitric oxide synthase; FBN1: fibrillin 1; HIF-1α: hypoxia inducible factor 1α; HSFs: human skin fibroblasts; HucMSCs: human umbilical cord mesenchymal stem cells; IKBKB: inhibitor of nuclear factor kappa-B kinase subunit beta; KLF6: Kruppel-like factor 6; miR: microRNA; Nrf2: nuclear factor erythroid 2-related factor 2; PRF: platelet-rich fibrin; SDF-1α: stromal-derived factor-1α; TNF: tumor necrosis factor; VEGF: vascular endothelial growth factor.
Further, an interaction between various genes and their interaction with miRNAs discussed above further potentiates the notion of targeting these molecular mediators to promote wound healing (Figure 6).
Interaction between genes and miRNAs discussed above as emerging targets to promote wound healing in diabetic foot ulcer (DFU). Networking done using https://www.networkanalyst.ca/NetworkAnalyst/.
The studies discussed above suggest that DFU pathogenesis and healing are a multifactorial process. Thus, targeting a single target or molecular process will not be enough to significantly promote wound healing in chronic non-healing DFUs. This indicates the need to target multiple therapeutic targets (Tables 4 and 5 and Figures 2 and 3), regulating various molecular processes involved in DFU wound healing. Most of the studies suggest that targeting inflammation to attenuate it and promoting angiogenesis and ECM remodeling should be the focus of future research to design a novel therapy with higher efficacy.
Gene therapy for DFUs offers high potential for treating non-healing, neuroischemic wounds, but faces significant barriers including high manufacturing costs, regulatory scrutiny on comparability, and the need for localized, stable, and durable delivery into hostile, infected chronic wound environments. Recently, the focus has shifted toward non-viral vectors and gene-activated matrices to overcome these limitations and improve clinical application. Gene therapies must meet stringent regulatory requirements regarding safety, efficacy, and consistency, often struggling to demonstrate long-term benefits in highly variable patient populations. Regulators require rigorous validation of production methods, particularly for customized or complex therapies, ensuring reproducibility and purity. Many promising therapies face slow progress in navigating the regulatory, evidentiary, and manufacturing complexities required for approval [148, 149]. Producing gene therapies, particularly autologous or complex ex vivo treatments, requires specialized facilities and faces significant logistical and cost hurdles. The need to transport and store, often time-sensitive, genetic materials creates bottlenecks, particularly for global distribution. To address scalability, the field is moving towards more standardized manufacturing processes to improve efficiency [150].
Hostile wound environment is another issue in gene therapy. The high protease and low vascularization environment of DFUs degrades growth factors, necessitating sustained, localized gene expression. Vector limitations are another issue. AAVs are commonly used but can face host immune responses, restricting repeat dosing, though non-viral vectors and matrices are improving this. Targeted delivery using novel approaches, such as gene activated matrix (GAM) technologies and non-viral vector systems (e.g., DNA-based), aim to maintain high local concentrations of therapeutic genes to enhance healing. While early studies showed promise in promoting wound closure, particularly in neuroischemic ulcers, consistent, long-term efficacy in diverse human DFU patients remains to be established on a wide scale [151].
High up-front cost is another issue with translating gene therapy to clinics. Gene therapies for DFUs are expected to be very costly, creating financial burdens on healthcare systems that require justification through high-value outcomes. Another concern is limited data for reimbursement. The limited long-term clinical data makes it difficult to secure reimbursement, which is crucial for widespread adoption. Long-term savings potential is another concern. While initial costs are high, successful gene therapies could potentially reduce the need for repeat treatments, hospitalization, and amputations, potentially leading to better cost-effectiveness in the long run [148, 152].
Gene therapy is poised to revolutionize medicine by shifting from treating symptoms to providing potential one-time cures for most diseases. In the context of DFU, gene therapy offers a promising, innovative future by targeting the root causes of chronic, non-healing wounds, such as poor angiogenesis, high infection rates, and impaired tissue regeneration. Novel approaches include topical, multi-target gene therapies AUP16 and HGF delivering VM202 for safer, more effective tissue repair. Targeted, non-viral, and viral vector-based approaches aim to accelerate angiogenesis, reduce inflammation, and promote tissue regeneration, with advanced therapies currently in phase III trials. Future research should focus on novel targets because targeting multiple molecular mechanisms will be beneficial in promoting wound healing in DFUs. The future of gene therapy for DFU should focus on overcoming the limitations of short-lived growth factor treatments by using genetic material to induce sustained, localized production of healing proteins. Future research should also focus on issues like clinical adoption of gene therapies facing significant hurdles, including prohibitively high costs, complex manufacturing logistics, and stringent long-term safety monitoring requirements. Regulatory challenges arise from inconsistent international standards and the need for, yet difficulty in, proving long-term, curative efficacy, leading to limited patient access even after approval.
AAV: adeno-associated virus
AGEs: advanced glycation end products
AI: artificial intelligence
Akt: protein kinase B
AMOs: anti-microRNA oligonucleotides
ANG1: angiopoietin 1
BM-MSCs: bone marrow-derived mesenchymal stem cells
circRNA: circular RNA
CSF1: colony-stimulating factor-1
CVD: cardiovascular disease
CXCR2: C-X-C motif chemokine receptor 2
DFU: diabetic foot ulcer
DM: diabetes mellitus
DN: diabetic neuropathy
DPP4: dipeptidyl peptidase 4
DUDT: diabetic under-the-skin/deep tissue
ECM: extracellular matrix
EPC: endothelial progenitor cell
EPS: extracellular polymeric substance
FGF: fibroblast growth factor
HGF: hepatocyte growth factor
IL: interleukin
LLLT: low-level laser therapy
LNPs: lipid nanoparticles
MD: mean difference
miRNAs: microRNAs
MMPs: matrix metalloproteinases
MSCs: mesenchymal stem cells
NLRP3: NLR family pyrin domain containing 3
Nrf2: nuclear factor erythroid 2-related factor 2
PAD: peripheral artery disease
PI3K: phosphoinositide 3-kinase
PRP: platelet-rich plasma
TGF: transforming growth factor
TNF: tumor necrosis factor
VEGF: vascular endothelial growth factor
IFA: Writing—original draft. VAG: Writing—original draft. VR: Conceptualization, Writing—original draft, Writing—review & editing. All authors read and approved the submitted version.
The authors declare that they have no conflicts of interest.
Insert content.
Not applicable.
Not applicable.
Not applicable.
The authors received no specific funding for this study.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 179
Download: 10
Times Cited: 0
