Review
Review

Affiliation:
1School of Pharmaceutical Sciences, Lovely Professional University, Phagwara 144411, Punjab, India
ORCID: https://orcid.org/0000-0001-6542-2464
Affiliation:
1School of Pharmaceutical Sciences, Lovely Professional University, Phagwara 144411, Punjab, India
ORCID: https://orcid.org/0000-0002-1969-6445
Affiliation:
2School of Bioengineering and Biosciences, Lovely Professional University, Phagwara 144411, Punjab, India
Email: yachanamishra@gmail.com
ORCID: https://orcid.org/0000-0002-3467-0518
Explor Immunol. 2025;5:1003212 DOI: https://doi.org/10.37349/ei.2025.1003212
Received: March 16, 2025 Accepted: August 05, 2025 Published: August 22, 2025
Academic Editor: Wenping Gong, The Eighth Medical Center of PLA General Hospital, China
The article belongs to the special issue Novel Vaccines development for Emerging, Acute, and Re-emerging Infectious Diseases
Messenger RNA (mRNA) vaccines represent a novel category of vaccinations with significant potential for the future. Recent studies have demonstrated the effectiveness of mRNA vaccines in combating various viral infections and cancer, particularly in cases where traditional vaccine platforms may not produce protective immune responses. In particular, mRNA vaccines have gained attention due to their quick development, scalable manufacturing, and ability to elicit strong immune responses. This review elucidates the synthesis of mRNA and mRNA vaccines, their mechanisms of action, and the strategies to enhance their delivery and address their advantages and limitations for viral disease. Many delivery strategies have been investigated in recent years, concentrating on nanoparticle-mediated mRNA vaccine delivery. The delivery mechanism is crucial for improving mRNA vaccine stability, biocompatibility, and targeting specific cells and tissues. By preventing mRNA degradation and increasing cellular uptake, nanocarriers significantly contribute to the stability and immunogenicity of mRNA vaccines. Nanoformulation functions not only as a carrier but also as a compartment that safeguards the mRNA from biological, chemical, and physical processes that may compromise its safety and efficacy. Despite these advances, challenges such as long-term safety and innate immune activation remain. Eventually, this review concentrated on future considerations necessary for the more efficient and safer deployment of mRNA, emphasizing the merits and drawbacks of the existing viral disease mRNA vaccines, with an eye toward future innovations and clinical applications.
One of the most successful public health initiatives, vaccination is recognized for its ability to control, eliminate, and even completely eradicate infectious diseases. Vaccines reduce the spread and severity of disease by preparing the immune system to identify and destroy pathogens [1]. Vaccines have saved countless lives and changed the face of global health since Edward Jenner’s groundbreaking research in 1796, which showed that exposure to cowpox could protect against smallpox [2]. Numerous vaccines for viral diseases are being employed for regular immunization, following significant improvements in the treatment and prevention of viral infections. Consequently, researchers are endeavoring to develop an efficacious vaccine for cancer. In 2006, the FDA sanctioned the first vaccine for cancer in human history, which inhibits human papillomavirus (HPV) 16 and 18 infections for over five years, therefore reducing the risk of cervical cancer [3, 4]. Despite substantial progress in vaccine development, many viral diseases continue to escape adaptive immune responses [5]. Moreover, the growing need for production at a large scale and swift advancement necessitates the creation of innovative vaccination strategies. Viral and non-viral diseases, such as human immunodeficiency virus (HIV) and cancer, require more research on vaccines to promote the development of innovative vaccine platforms.
The messenger RNA (mRNA) vaccination represents a novel approach that integrates immunology and molecular biology. The somatic cells are introduced with antigen-encoded foreign mRNA to facilitate antigen synthesis via the expression system [6]. These synthetic antigens can produce an immunological response [7]. In 1990, researchers introduced mRNAs into mouse somatic cells in vivo by utilizing mRNA expression vectors, resulting in the production of chloramphenicol acetyltransferase, luciferase, and beta-galactosidase [8]. In 1992, Jirikowski et al. [9] administered vasopressin and oxytocin-encoded mRNA in genetically mutated mice with diabetes insipidus partially alleviated the condition. Despite significant discoveries thereafter, no meaningful advancements in mRNA research occurred due to challenges such as amplified immunogenicity, instability, and the absence of an adequate delivery method for mRNA vaccines [10–12].
Over the past decades, advancements in research and experimental methodologies have enhanced the effectiveness, safety, and large-scale industrial manufacturing of mRNA vaccines [13]. The mRNA vector lacks traits such as genomic integration, robust immunogenic responses, and antibiotic resistance [14–16]. Furthermore, nucleases swiftly decompose single-stranded RNA (ssRNA) [17]. While the damaged mRNA mechanisms provoke excessive activation of the immune system, the development of a safe and efficient delivery method utilizing modified mRNA can mitigate negative effects and improve efficacy [18, 19]. Enhanced therapeutic effectiveness can be achieved by mRNA carriers and modified mRNAs. For example, a herpes simplex virus 2 nucleoside-modified mRNA vaccination reduced viral levels in infected individuals [20]. The liposome conjugated with mannose has been used to transport the mRNA into cells. The carrier vector not only enhanced mRNA overexpression but also safeguarded mRNA against degradation by upregulating the mannose receptor on cell membranes [21]. Modified mRNAs and numerous delivery vectors have been extensively examined for their therapeutic application, particularly during the COVID-19 pandemic. Approved mRNA vaccines, notably for SARS-CoV-2 (severe acute respiratory syndrome coronavirus 2), show strong protection against severe illness, with others in development for viral diseases, especially for immunosuppressed groups. Their adaptability and precision make mRNA vaccines promising for future widespread use [22]. The mass production of mRNA vaccines depends on translational research, which is essential for expediting the speed of manufacturing. The in vitro translational research approach swiftly identifies structures and formulations in both clinical and preclinical research [23].
Notwithstanding these achievements, there are still significant gaps in the literature. Comparing the delivery methods, effectiveness results, and mechanistic insights of various mRNA vaccine applications for various viral diseases, including Zika, rabies, influenza, COVID-19, and HIV, is not well done. This review addresses these gaps by examining the synthesis and mechanism of mRNA vaccines, evaluating delivery technologies—especially nanoparticle-mediated systems—and analyzing their application across several high-priority viral diseases. This review aims to critically assess both the bottlenecks and breakthroughs in mRNA vaccine development and delivery to understand future research directions and improve global preparedness for emerging infectious threats.
The preclinical evidence facilitates future clinical assessment. The fast production of mRNA vaccines positions them as a viable therapeutic approach. mRNA vaccines signify a notable shift from conventional vaccination technology, presenting certain benefits but also encountering new obstacles. To compare mRNA vaccines with traditional vaccine platforms, including live-attenuated, inactivated, subunit, and viral vector vaccines, it is crucial to analyze fundamental attributes such as production techniques, immune responses, safety profiles, and storage necessities.
Live-attenuated vaccines: Live-attenuated vaccinations utilize a weakened version of the virus or bacterium that may multiply within the body without inducing sickness. Examples include the measles, mumps, and rubella (MMR) vaccination, as well as the yellow fever vaccine. Live-attenuated vaccines often elicit robust humoral (antibody) and cellular (T cell) immune responses, resulting in enduring protection with a reduced number of doses. In several instances, a solitary dose is adequate for enduring protection (e.g., the yellow fever vaccination). A minor threat exists that the attenuated pathogen may induce illness, particularly in immunocompromised persons (e.g., those with HIV or cancer). These vaccines often require refrigeration for storage, and some types may exhibit reduced stability compared to inactivated vaccines [24–26].
Inactivated vaccines: Inactivated vaccinations utilize viruses or bacteria that have been rendered nonviable, preventing replication. Examples are the inactivated poliovirus vaccine (IPV) and the hepatitis A vaccine. Inactivated vaccinations are regarded as highly safe as they lack live germs, thereby eliminating the danger of disease manifestation. They exhibit greater stability at elevated temperatures compared to live-attenuated vaccines and often do not need ultra-cold storage. Inactivated vaccinations often elicit a less robust immune response compared to live-attenuated vaccines, frequently necessitating several doses or booster injections. These vaccines often elicit a vigorous antibody response but may not provoke a significant T-cell response, which is crucial for enduring protection [27, 28].
Subunit vaccines: Subunit vaccines are pure components of the pathogen, including proteins or carbohydrates, which produce an immune response. Examples include the HPV vaccination and the Haemophilus influenzae type b vaccine. Subunit vaccinations, including only components of the pathogen, pose no risk of infection, rendering them safe for immunocompromised persons. These vaccines are generally more stable than live-attenuated vaccinations and may be stored under regular refrigeration conditions. Subunit vaccinations often produce a less robust immune response than live-attenuated vaccines and may need adjuvants or several doses to improve efficacy. These vaccinations frequently induce only an antibody response and may not produce the complete spectrum of immunological protection (e.g., T-cell responses) [29, 30].
Viral vector vaccines: Viral vector vaccines such as the AstraZeneca COVID-19 vaccine and the Ebola vaccine utilize a distinct virus (unrelated to the target disease) to introduce genetic material into cells, prompting the production of the pathogen’s proteins to produce an immune response. Similar to live-attenuated vaccinations, viral vector vaccines can produce both humoral and cellular immune responses, potentially resulting in prolonged protection. Like mRNA vaccines, viral vector vaccines do not need the cultivation of the target pathogen, facilitating expedited development and manufacture. Certain people may possess pre-existing immunity to the viral vector employed, thereby diminishing the vaccine’s efficacy. While not as rigorous as mRNA vaccines, viral vector vaccines sometimes need refrigeration, posing difficulties in resource-constrained environments [31, 32].
Every vaccination platform possesses unique benefits and drawbacks. mRNA vaccines are notable for their rapid creation and adaptability; yet, issues related to stability and cold-chain storage persist. Live-attenuated vaccinations provide robust protection but entail a little risk of inducing illness. Inactivated vaccinations are safer; nonetheless, they generally need several doses. Subunit vaccines have good safety and stability, although they may need adjuvants to elicit adequate protection. Ultimately, viral vector vaccines produce a robust immune response; however, they may be influenced by pre-existing immunity to the vector. Comprehending these distinctions is essential for choosing the appropriate vaccination platform for certain illnesses and demographics. The mRNA vaccines can be designed and manufactured more rapidly than traditional vaccines, as they do not require the growth of viruses or bacteria (e.g., for live-attenuated or inactivated vaccines) [33]. The mRNA platform can be quickly adapted to target new variants of a virus, making it highly responsive to emerging infectious diseases [34].
A thorough narrative overview of previous studies on mRNA vaccines, their role in vaccine production, and their efficacy in preventing viral infections is presented in this review. It examines the development of vaccination technologies by contrasting mRNA-based platforms with traditional techniques, including live-attenuated, inactivated, and subunit vaccines, etc. The review focuses especially on mRNA vaccine delivery systems, describing their advantages, present problems, and possible fixes to several viral illnesses. Additionally, it draws on data from clinical trials and meta-analyses to highlight the progress of mRNA vaccines across various stages of development, supported by tables summarizing trials using diverse delivery strategies.
A thorough literature review was conducted using major databases such as PubMed and ClinicalTrials.gov to uncover relevant publications and clinical trials. Both single and multiple combinations of keywords like “mRNA vaccine”, “mRNA synthesis”, “mRNA vaccine formulation”, and “mRNA delivery”, as well as disease-specific terms like “mRNA vaccine for COVID-19”, “Zika virus”, and “HIV”, were employed. The procedures involved in mRNA production, formulation, and the variety of delivery devices used are examined in this narrative review. Clinical trials, their advancement, and the relative benefits of mRNA vaccines over traditional vaccine platforms in the treatment of viral infections are all summarized. Additionally, the review addresses the application of mRNA vaccines in viral disease, presents challenges in the field, and proposes possible solutions.
Synthetic mRNA is generated through the transcription of a complementary DNA (cDNA) template, usually plasmid DNA (pDNA), in vitro with the use of bacteriophage RNA polymerase [35]. The initial phase of this process involves the synthesis of pDNA. While mRNA synthesis appears more complex than pDNA production, raw pDNA often contains bacterial DNA remnants and exists in various forms (supercoiled, relaxed, or linear), complicating the consistent generation of pure and stable pDNA for applications such as vaccination. Linearization of pDNA results in the removal of bacterial DNA during transcription by RNA polymerase [36].
The synthetic mRNA produced contains an open reading frame (ORF) surrounded by essential components: a 5' cap (7-methyl-guanosine) and a 3' poly(A) tail [37, 38]. The pDNA utilized for transcription must contain a bacteriophage promoter, an ORF, and a restriction site for linearization. Transcription is facilitated by RNA polymerase (T7, T3, or SP6) along with the required nucleotides. The production of capped mRNA can be achieved by incorporating a cap analog such as m7G(5')-ppp-(5')G [39]. Following transcription, DNase treatment eliminates the pDNA template and bacterial DNA [40].
The stability of mRNA is crucial for facilitating effective protein expression. Alterations to the 5' UTR (untranslated region) and 3' UTR surrounding the ORF can enhance mRNA stability and expression levels significantly [41]. A 5' cap, exemplified by the LNA (locked nucleic acid)-modified cap analog (m7(LNA)G[5']ppp[5']G), enhances stability and translation efficiency [42, 43]. Enzymatically added caps demonstrate greater efficacy compared to synthetic analogs [44, 45]. The poly(A) tail is essential for mRNA stability; its removal leads to destabilization, decreased translation efficiency, and impaired polysome formation [46, 47].
Furthermore, the incorporation of modified or synonymous codons in the mRNA sequence may enhance stability through modifications to the secondary structure or by improving translation fidelity [48, 49]. Elevating the G:C content enhances stability [50]. Modifying elements such as codons, 5' UTR, 3' UTR, cap, and poly(A) tail enhances mRNA stability and promotes efficient immune responses [51, 52].
mRNA vaccines offer advantages like rapid development, scalability, and production efficiency. Their manufacturing process includes three stages: synthesis, purification, and formulation of the mRNA component. This section discusses the recent advances in each phase to enhance vaccine production.
In the synthesis phase, mRNA is produced via in vitro transcription using RNA polymerase enzymes (T7, SP6, or T3) to transcribe the gene of interest from a linearized DNA template [53]. Once synthesized, the mRNA is purified to remove contaminants like residual nucleotides, enzymes, and DNA. Standard methods like DNase digestion followed by lithium chloride precipitation are commonly used, but they may not fully eliminate impurities such as double-stranded RNA or incomplete mRNA transcripts [54]. Ineffective purification can lead to poor translation efficiency and altered immunogenicity. Advanced techniques like reverse-phase HPLC have improved purification, leading to higher transfection and protein synthesis [55]. Chromatography methods such as size exclusion and ion-pair reverse-phase chromatography are often employed. Ion-pair reverse-phase chromatography is particularly effective in isolating mRNA from contaminants due to charge differences, offering higher yields and purity compared to size exclusion [56–59]. Other methods, like affinity chromatography using oligo dT beads and tangential flow filtration, are also used for further refinement [60, 61].
In the formulation phase, mRNA is encapsulated in lipid nanoparticles (LNPs) to prevent degradation and enhance delivery. LNPs, which are FDA-approved for mRNA vaccines, are formed by mixing lipids [ionizable lipids, cholesterol, PEG (polyethylene glycol)] in an organic solvent with mRNA in an aqueous buffer. This creates electrostatic interactions, forming stable nanoparticles. After synthesis, the solution undergoes dialysis to remove solvents and adjust pH. Microfluidic mixing is the preferred method for creating uniform LNPs with high mRNA encapsulation efficiency. Platforms like NanoAssemblr® are widely used for precise and consistent nanoparticle formation at both research and commercial scales [62, 63].
The mechanism of action of mRNA vaccines involves the introduction of synthetic mRNA into host cells. This mRNA encodes a specific antigen, which is then translated by the host’s ribosomes. The resulting protein is recognized by the immune system, prompting an immune response that includes the production of antibodies and the activation of T cells. This process ultimately prepares the immune system to recognize and combat the actual pathogen upon exposure.
mRNA serves as a messenger transcribed from genomic DNA within the nucleus and translated by ribosomes in the cytoplasm to synthesize proteins. mRNA is initially transcribed from DNA as primary mRNA through the action of RNA polymerase. This mRNA comprises exons and introns. It undergoes additional processing, including intron removal, the addition of a 5' cap, and a poly(A) tail at the 3' end, to form mature mRNA [64, 65].
In mRNA vaccination, the mRNA is synthesized externally and subsequently delivered to cells, where it penetrates the cell membrane for translation into the target protein within the cytoplasm. The vaccine-derived mRNA, akin to natural mRNA, binds to ribosomes for the process of translation. In the process of translation, the poly(A) tail interacts with poly-A-binding proteins (PABP), while eukaryotic initiation factors are associated with the 5' UTR cap. The ribosome decodes the codons in the mRNA coding region, translating them into amino acids [66]. The mRNA in vaccines comprises a 5' cap, 5' UTR, coding region, 3' UTR, and poly(A) tail, all essential for effective translation and antigen production [67].
The efficacy of mRNA vaccines is significantly dependent on the activation of the immune system; however, the role of type I interferon (IFN) is still a subject of debate. Inhibition of type I IFN receptors has been demonstrated to enhance immune responses, leading to increased production of antigen-specific antibodies and CD8+ T cells [68]. Factors that influence the immune response encompass the half-life of the antigen, the presentation of antigens on MHC (major histocompatibility complex) class I and II molecules, and interactions with components of the innate immune system, including NK cells and macrophages.
An important benefit of mRNA vaccines compared to pDNA vaccines is that mRNA does not necessitate nuclear entry or transcription for the synthesis of the target protein [69]. mRNA delivery encounters significant challenges, especially in traversing the cell membrane and evading endosomal entrapment [70]. The mRNA encodes the antigen, initiating an immune response that enables the body to identify and fight the pathogen upon subsequent exposure [64].
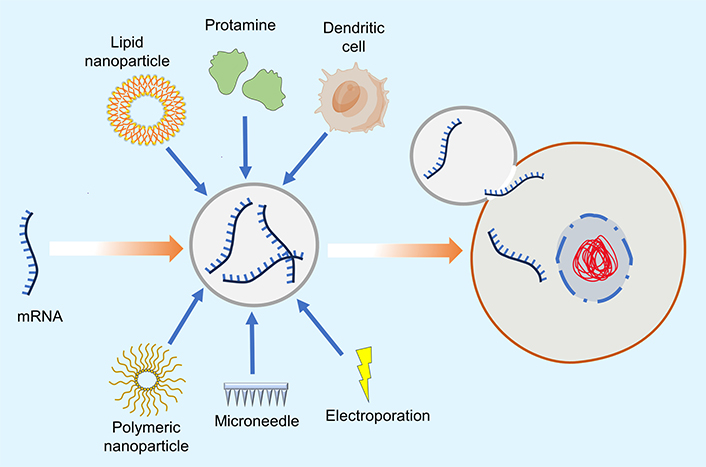
Various methods used in mRNA delivery are represented in Figure 1.
Preliminary research has shown that in vivo nude mRNA delivery can produce an immunotherapeutic reaction in mice [8]. Currently, administration methods for mRNA encompass subcutaneous, intradermal, intranodular, intramuscular, intravenous, and intratumoral injections, which are vital for stimulating antigens and initiating a response from the immune system [33, 71, 72]. Phua et al. [73] found that subcutaneous nude mRNA injection in mice surpassed the efficacy of nanoparticle-based delivery of mRNA. Lint et al. [74] discovered that when mRNA is directly injected into the tumor through intratumoral injection, it provokes a suitable response from the immune system and is considered a potentially feasible vaccine delivery technique in the coming years. Currently, the direct injection of unencapsulated mRNA is mostly employed for the treatment or prevention of viral infections [75]. Nonetheless, despite the potential to produce a response from the immune system, the efficacy of this transport route is comparatively limited, and nude mRNA is frequently subject to fast degradation post-injection. The direct injection of nude mRNA is overly simplistic and rudimentary for application in human patients; it is frequently utilized as a method to administer modified mRNA vaccines in conjunction with alternative delivery techniques to enhance vaccination efficacy.
The efficacy of antigen release by mRNA is enhanced using many established physical techniques, including microneedles, electroporation, and gene gun, among others. Electroporation possesses an adjuvant effect, capable of recruiting pro-inflammatory cells and stimulating cytokine synthesis at the site of administration, hence enhancing the immune response of the mRNA vaccine [76]. It also enhances mRNA transport effectiveness without requiring additional receptor modalities, minimizing unwanted immunological responses [77]. Callis et al. [78] discovered that transfection efficacy in plant and animal cells can be increased by the electroporation of the mRNA. In 2017, the electroporation approach increased the mRNA transfection efficacy (50–90%) in dendritic cells (DCs) [79]. The gene gun technique employs pressurized He gas as an accelerating force to propel gold particles coated with mRNA into host cells, serving as an effective way for the delivery of mRNA [80]. Qiu et al. [80] employed the gene gun technique on mouse skin for the introduction of human alpha-1 antitrypsin mRNA, effectively inducing an antibody release. Peking et al. [81] treated hereditary skin disorders by mRNA-based formulation, with mRNA successfully delivered to mice targeted skin layer using the gene gun method. However, the gene gun technique is hardly employed in humans or large animals. Physical methods of delivery of mRNA may influence the function of cells and their physiological structure and potentially result in aberrant cell death. Consequently, using physical mRNA delivery in humans poses significant risks [33, 76].
DCs are among the most powerful immune system stimulants for antibody release. DCs use the MHC for the delivery of processed antigen to CD4+ and CD8+ T cells, hence initiating cellular immune response [33, 82]. Concurrently, DCs can deliver antigens to B cells, triggering a humoral immune response [83]. The delivery of mRNA involves transfecting mRNAs that encode proteins, peptides, or other immune stimulants in DCs, followed by the host organism being infected with DCs carrying mRNA to initiate the specific response from the immune system based on the antigen [84]. This method operates independently of additional carrier molecules and achieves great delivery efficiency. It is extensively utilized in clinical research, animal models, and pre-clinical trials [85–88]. This method has mostly been utilized in the treatment of cancer due to the predominance of eliciting cellular immune responses [89]. Nonetheless, the mRNA transfection rate is rather poor when relying just on DC endocytosis, prompting the frequent utilization of electroporation to enhance the rate of transfection of the mRNA [90]. Gay et al. [90] employed electroporation to DCs carrying HIV-specific antigens encoded in mRNA for HIV therapy. Followed by at least twice intradermal injection, cytotoxic T-lymphocytes (CTLs) CD28+/CD45RA–CD8+ immune responses were induced in HIV patients. A significant obstacle to the practical implementation of DC-loaded mRNA is the time and cost-intensive manufacturing procedure, which limits the scalability of DC-based mRNA vaccines. Furthermore, within hours of transfection, the immune system produced a diminished response, resulting in a decreased therapeutic response [91]. Considering these factors, diseases necessitating substantial mRNA vaccine administration in the immediate future should prioritize delivery methods with rapid manufacturing capabilities.
The polymer-based delivery system uses a cationic polymer as a vector for mRNA delivery [92, 93]. Frequently utilized polymer delivery polymers are poly(beta-amino esters) (PBAEs) and polyethylenimine (PEI), which are a few examples. PEI is the most extensively utilized polymer. It is a water-soluble cationic polymer recognized by branched, linear, or dendritic structures, commonly used for mRNA transfection [94, 95]. Linear PEI is marketed as jetPEITM was previously utilized for siRNA (short interfering RNA) and DNA transfection and can be used for transfection of mRNA [92, 96]. Nonetheless, PEI possesses inherent cytotoxicity that is resistant to degradation; hence, researchers frequently alter low-molecular-weight PEI with fatty chains to mitigate its toxicity [94, 97, 98]. PBAEs are biodegradable polymers initially created for DNA transfection [99]. Kaczmarek et al. [100] created a delivery system based on a polymer by combining lipid-PEG with PBAEs, achieving high effectiveness and stable mRNA transport, after successfully injecting intravenously. In 2007, research showed that PBAEs can facilitate mRNA delivery, achieving elevated mRNA transfection levels in vitro in the absence of serum proteins [101]. The ease of synthesis has led to the creation of numerous PBAEs, some of which improve in vivo mRNA stability [92, 102, 103]. Lipids and PBAEs can be combined to enhance the stability of mRNA in blood. Compared to lipids, polymer materials are strong contenders for mRNA transport. However, the cytotoxicity of polymer complexes, similar to cationic lipids, has hindered wider application [104]. In addition to modifying polymer-based vectors with other materials to enhance their capabilities, optimizing branching patterns and molecular weight appears to be a promising approach.
LNPs are the sole nano-delivery technology for mRNA vaccines that are clinically successful and have received approval for human use. The Pfizer and Moderna SARS-CoV-2 mRNA vaccination uses LNPs to transport the mRNA to the target. Cationic liposomes are frequently utilized as RNA carriers, since lipids are positively charged and RNA is negatively charged, which creates electrostatic complexes, hence enhancing mRNA transport efficacy [105]. The mRNA-lipid combination, along with other excipients, may produce LNP ranging from 80 to 200 nm, which can reach the cell cytoplasm through endocytosis. Their most common composition comprises cholesterol, natural phospholipids, ionizable cationic lipids, and PEG [76]. The mRNA is self-aggregated due to cationic lipids forming approximately 100 nm particles and, through endocytosis, reaches the cytoplasm, gets ionized, and releases mRNA. Cholesterol stabilizes the LNP, PEG increases its half-life, and phospholipids help form the lipid bilayer [33, 92]. LNP encapsulates the mRNA in the core, which safeguards it from degradation. The lipophilic characteristics of the LNP component facilitate the transport of mRNA lipid complex across the membrane of the cell through endocytosis [76, 91]. LNP is frequently utilized as a delivery method for siRNA in initial studies [106].
Currently, LNP is extensively utilized in the delivery of mRNA. Geall et al. [107] employed an mRNA-lipid complex to administer a self-amplifying RNA, resulting in elevated mRNA expression levels when compared to naked mRNA in mice, while also efficiently producing CD4+ and CD8+ T cell immunological responses. Various delivery routes can elicit distinct immune stimulation responses from mRNA LNPs, potentially fulfilling the therapeutic requirements for diverse illnesses. Pardi et al. [108] discovered that administering mRNA LNPs at the correct dosage via intradermal, intramuscular, and subcutaneous routes might facilitate localized gene translation. Common mRNA-LNP delivery methods are tracheal inhalation, intraperitoneal injection, and intravenous injection, the latter showing the highest systemic delivery efficiency [109]. However, the methods by which mRNA escapes from electrostatic complexes to achieve a free state for functionality in the cytoplasm remain inadequately comprehended. The in vivo ambient pH alters the ionization state of lipid-carrying mRNA is considered crucial to the release mechanism [104]. Simultaneously, more investigation is needed into decreasing the toxicity, and the management of immunogenicity in cationic lipid-based delivery systems is critically required.
Cellular transport and encapsulation can be enhanced by using phospholipids in LNP formulation. To extend the circulation duration for LNP, the cholesterol level should be increased while decreasing the level of phospholipid. Some studies indicate that higher phospholipid molar ratios enhance LNP delivery efficiency. Zwitterionic phospholipids may help stabilize electrostatic interactions between mRNA, lipids, and surrounding water molecules. However, the precise role of phospholipids in mRNA transport through LNPs remains unclear [110].
Protamines are positively charged proteins with resinous structures. The combination of protamine with mRNA in various ratios of mass can produce electrostatic mRNA-protamine complex nanoparticles of various sizes [111]. This compact conjugate structure may efficiently safeguard mRNA from degradation by RNase enzyme, and this complex can produce a robust immunological response from immune cells, including DCs, neutrophils, NK cells, B cells, and monocytes [112–114]. It suggests that protamine may serve as both an mRNA transporter and an immunological activator. In 1961, protamine was among the initial materials investigated for the delivery of RNA [115].
Fotin-Mleczek et al. [116] administered a vaccine that has protamine as a carrying agent of an mRNA to the tumor, effectively inducing a comprehensive anti-tumor-specific response. At the mass ratio of mRNA to protamine of 2:1, the resultant complex measures around 300 nm, exhibiting relative stability and eliciting robust immunological activation alongside elevated cytokine levels, but with a considerable inhibition of protein production. Consequently, it is widely believed that while mRNA-protamine complexes induce immune responses, translation efficiency may be limited [116, 117]. In recent years, mRNA-protamine formulations have been extensively utilized in clinical studies, demonstrating significant therapeutic efficacy in conditions such as rabies [110].
Currently, over 30 diseases globally can be averted with vaccination. Despite decades of rigorous endeavors to create effective vaccines for formidable viruses that induce recurrent or chronic infections, such as HIV and respiratory syncytial virus (RSV), success has not yet been achieved. Moreover, the major challenge for emerging virus vaccines is the urgent necessity for swift development, which conventional methods cannot adequately address, as demonstrated by the outbreaks of Zika virus, Ebola, and Middle East Respiratory Syndrome (MERS) in the last decade, as well as the outbreak of COVID-19 between 2020–2022 [118]. Therefore, a more powerful and adaptable platform for vaccine delivery must be created as it is necessary.
In 1990, the in vivo use of in vitro transcribed (IVT) mRNA was documented, demonstrating protein synthesis from directly injected reporter gene mRNA into the muscle of the mouse [8]. Nonetheless, apprehensions over mRNA instability, robust innate immune responses, and inadequate in vivo transport efficiency led to a lack of significant investment in the development of mRNA vaccines [33, 119].
Leveraging significant technology advancements and research investments over the last few years, mRNA has appeared as a viable platform for vaccines for various viral diseases. The mRNA vaccine often provides various benefits over traditional methods, such as protein subunit, inactivated viral, live attenuated, and genetic material-based vaccines. Primarily, mRNA vaccination is safe, being a non-integrating platform or non-infectious, it poses no possible danger of mutagenesis or infection. Secondly, high efficacy of mRNA vaccination, nucleoside modifications significantly enhance mRNA stability and translational efficiency, while LNPs serve as effective carriers for in vivo mRNA delivery, facilitating immediate acceptance and expression in targeted cells, ultimately resulting in robust and adaptive cellular and humoral response from the immune system [118, 120]. Thirdly, the rapid development and adaptability of mRNA vaccines may be achieved within a few days using acquired gene sequence data. This method is very adaptable and applicable to nearly all protein targets [121]. Table 1 shows a SWOT analysis of mRNA for viral disease.
SWOT analysis of mRNA vaccines in the context of viral diseases
| SWOT analysis | Internal factor* | |
|---|---|---|
| External factor# | Strengths (S)
| Weaknesses (W)
|
Opportunities (O)
| OS strategy
| OW strategy
|
Threats (T)
| TS strategy
| TW
|
* Internal factors: attributes inherent to mRNA vaccine technology or design (e.g., scalability, immunogenicity). # External factors: influences from the regulatory, social, geopolitical, or scientific situation (e.g., distribution logistics, mutations, public perception). mRNA: messenger RNA
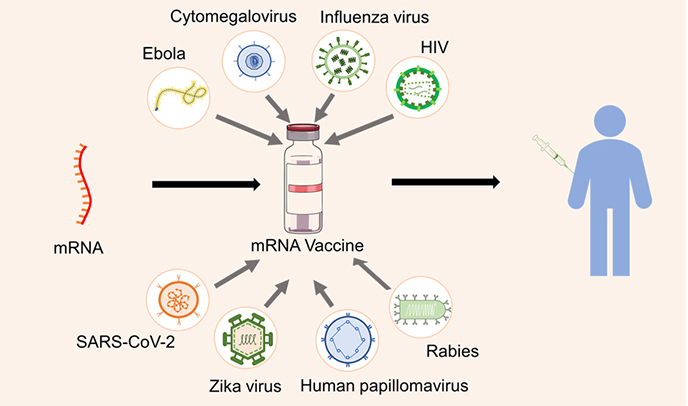
Various viral diseases prevented using mRNA vaccines are shown in the Graphical abstract.
Influenza virus vaccines have been utilized for over 80 years, although they remain a significant focus of investigation-based study and a considerable demand on community health resources due to the necessity for frequent updates to address the continually evolving viruses [122]. The inaugural influenza vaccine was employed in the mid-1930s, following the recent discovery of influenza A viruses (IAVs) a few years earlier at Mill Hill. Influenza immunization originally entailed subcutaneous administration of formalin-inactivated viruses cultured in murine lungs; subsequently, egg-cultivated viruses that are inactivated were created by Jonas Salk and Thomas Francis Jr. for the military of the USA. In the following decades, both IAVs and IBVs were encompassed in influenza vaccination and are now generally advocated for yearly administration. These vaccinations are crucial instruments for managing seasonal influenza and addressing influenza pandemics [123–125].
Notwithstanding the available vaccine alternatives, viruses of seasonal influenza are projected to result in more than a few million infections and are linked to around 650,000 fatalities annually worldwide [126, 127]. Vaccine policy and coverage differ significantly among nations: in 2019, immunization rates for those aged 65 and older ranged from around 6% to over 70% of the population. In nations with substantial vaccination coverage, the virus of seasonal influenza exerts a significant illness burden, which can be largely ascribed to the deficiencies of existing vaccines [126, 128].
A limited number of pre-clinical and early clinical experiments for mRNA-based influenza vaccines have been performed or are underway. A phase I experiment was conducted to assess the safety of mRNA vaccines carried by liposomes containing H7N9 or H10N8, the full-length IAV, avian viruses with pandemic potential, can be delivered either intradermally or intramuscularly [129]. The immunizations were well handled and produced elevated, enduring antibody titers, comparable to current vaccines. The adverse effects documented in the 2019 research, including discomfort and edema at the site of administration, myalgia, and exhaustion, are analogous to those associated with mRNA vaccines for coronavirus [130]. The participants are recruited for trials by Moderna to evaluate a quadrivalent seasonal formulation of mRNA-1010, and preliminary findings after its first phase clinical trial in old and adults indicate it is safe and generates antibody responses [34]. The phase II clinical study performed by Pfizer of mRNA vaccines targeting seasonal influenza in people above 65 years [131]. Multiple firms are exploring the feasibility of a multivalent vaccination incorporating antigens from both the influenza virus and influenza and SARS-CoV-2. ‘Novavax’ is a nanocarrier-based vaccine, while Moderna has also declared to merge SARS-CoV-2 and influenza antigens into a unified vaccine [132]. Although influenza mRNA vaccines have been under research for some years, the efficacy of mRNA vaccines against SARS-CoV-2 has elevated their clinical application to a prominent prospect. The ability to manufacture substantial volumes of tailored, precisely regulated vaccinations in a comparatively short timeframe is appealing in relation to existing vaccine platforms. The regulatory inquiries and logistical challenges associated with the implementation of novel technology for the deployment of coronavirus vaccines which is expected to facilitate the introduction of next-generation influenza vaccines soon [133]. There is growing interest and investment in mRNA-based influenza vaccines, with several candidates in or completing phase III trials. It’s also crucial to monitor efforts like the WHO/MPP mRNA Technology Transfer Programme to track how low and middle-income countries are advancing in influenza vaccine development [134].
The initial human case of Zika virus infection was identified in 1952 in Nigeria. It is spread by Aedes spp. mosquitoes, remained inconspicuous for years until epidemics transpired from 2007 to 2017 in America, French Polynesia, and Yap Island. After the 2015 Brazil epidemic, which marked the initial instance of the indigenous Zika virus spread, initiatives started to emphasize the Zika virus investigation and the formulation of vaccines for the Zika virus. The worldwide concern for pregnant women stemmed mostly from the adverse effects on fetuses infected with the Zika virus in Brazil and other nations globally [135]. Neutralizing antibodies are essential agents of shield against infections and have been associated with the effectiveness of vaccinations [75, 136, 137]. Despite global interest in Zika resulting in several vaccine candidates, there is still no approved vaccine for the disease.
Despite the extensive range of vaccine platforms for the Zika virus, only two mRNA technology-based vaccine candidates have undergone clinical testing. The mRNA vaccination technology is a secure and efficacious method for developing protective immune responses. In contrast to existing vaccination candidate platforms, mRNA vaccines eliminate the danger of insertional and infection mutagenesis, while also circumventing the potential for antivector protection, so permitting recurrent delivery [33]. The effectiveness of an mRNA vaccine candidate in an AG129 murine model. A single administration of Zika virus prM-E mRNA-LNP conferred protection to rats following a deadly Zika virus infection. In comparison to the placebo group, vaccinated animals exhibited no clinical symptoms or weight reduction and had decreased virus loads. Finally, the BALB/c mice vaccination, accompanied by the isolated splenocytes analysis for T cells, revealed specific antigens for CD4+ and CD8+ T-cell responses. This study facilitates the usage of liposomes for mRNA vaccine delivery by liposome [138].
Since the initial clinical identification of acquired immunodeficiency syndrome (AIDS) and the subsequent discovery of the retrovirus HIV in the early 1980s, the HIV/AIDS epidemic remains a significant global health issue despite four decades of rigorous study. In 2019, there were 1.7 million new infections and 690,000 AIDS-related fatalities worldwide; in 2020, 38 million individuals were living with HIV. Contemporary prevention and treatment modalities for HIV/AIDS encompass the administration of antiretroviral medications for pre-exposure prophylaxis (PrEP) and antiretroviral therapy (ART), which have facilitated the transition of AIDS from a life-threatening condition to a manageable chronic disease [139]. Nonetheless, the drugs are costly, need rigorous compliance with dose protocols for effectiveness, and induce adverse effects. Furthermore, many HIV-infected patients acquire treatment resistance, necessitating alterations in their pharmaceutical regimen. Access to treatment is a significant obstacle, especially in low- and middle-income countries, as indicated by the consistently elevated rates of new infections over the past decade [140]. Consequently, preventive vaccination is a fundamental element of a comprehensive approach to eradicating the pandemic. Creating an effective HIV vaccine has, however, been shown to be exceptionally challenging. As the pandemic approaches its fifth decade, there are no authorized vaccines for HIV, with just one promising clinical research (RV144 trial) exhibiting a low effectiveness of 31% [141].
The mRNA vaccine utilizes mRNA that encodes an immunogen, which is converted into protein once delivered to host cells. In 1989, it was discovered that mRNA contained in cationic liposomes could transfect mouse cell lines [142]. The protein expression and translation were shown following the injection of nude mRNA into the skeletal muscle of the mouse [8, 143]. Nonetheless, employing mRNA for vaccines was impractical due to the instability of RNA molecules, insufficient delivery mechanisms, challenges in the large-scale production of mRNA, and unregulated activation of innate immunity via RNA sensors [143, 144]. Current technical advancements and enhanced delivery systems have mitigated these challenges, leading to the emergence of mRNA vaccines as a potential platform for antigen delivery. The mRNA-based vaccines provide several advantages compared to conventional vaccination platforms, such as enhanced safety, effectiveness, and streamlined manufacturing processes. The absence of effective delivery mechanisms for molecules of mRNA was a significant impediment to the advancement of mRNA vaccines. The optimal delivery mechanism will enhance cellular uptake with minimal toxicity and safeguard mRNA against destruction [145].
Rabies is a deadly neurological illness caused by the neurotropic rabies virus, affecting infected animals and humans. It is an ssRNA virus belonging to the Lyssavirus family, transmitted by several animal vectors; dog bites are the cause of more than 95% of rabies cases in humans, while additional carriers include bats, skunks, raccoons, and coyotes [146]. The true impact of rabies is challenging to assess due to case under-reporting and insufficient monitoring data; nonetheless, the risk of exposure is estimated to be more than 3 billion people worldwide, with 59,000 fatalities annually attributed to canine rabies. Regrettably, rabies continues to claim numerous lives, despite Louis Pasteur’s introduction of the first effective human vaccination against the disease over 130 years ago [147, 148].
The application of mRNA technology for the efficient synthesis of mRNA that is precisely tailored to the encoded antigen offers several benefits compared to current vaccination technologies in the creation and manufacture of preventive vaccines for infectious diseases. The mRNA vaccines do not incorporate any living viral components, unlike attenuated or live vaccinations, hence eliminating the risk of consequent infection and reversion to pathogenicity [118]. The prophylactic mRNA vaccine, which was first developed for rabies, has been advanced through in vivo animal models before human evaluations. The choice to prioritize rabies was influenced by many considerations. The rabies genome is well-defined and uncomplicated, as is the immune response target, the RABV-G protein antigen [149]. Numerous licensed vaccinations are available and established standardized assays for assessing the RABV-G antigen immune response, allowing for straightforward comparisons of rabies mRNA vaccine candidates with licensed and commercially available rabies vaccines [150].
Recent reports have detailed a technological framework for the versatile and precise synthesis of protein antigens in mRNA, which has been utilized for rabies. The investigations at the preclinical level have shown that intradermal or intramuscular administration of mRNA can produce cellular and humoral immune responses against the protein antigen [151].
COVID-19 is an illness induced by the SARS-CoV-2, which is a positive ssRNA virus enveloped by protein. It belongs to the Betacoronavirus genus within the Coronaviridae family. The whole genome comprised 29,881 nucleotides in the ancestral SARS-CoV-2 strain, with a 3' poly(A) tail and methylated 5' cap, encoding 9,860 amino acids that represent 4 structural proteins, 9 accessory proteins, and 16 nonstructural proteins. More than 220 countries have been devastated by COVID-19, resulting in more than 400 million incidence cases and 5.75 million fatalities [152–154].
The initial group of vaccines for COVID-19 is mRNA vaccines undergoing clinical testing. According to the WHO report, the number of COVID-19 vaccines exceeded 300, including 47 mRNA vaccines [155]. Moderna (mRNA-1273), Pfizer-BioNTech (BNT162b2), and CureVac mRNA vaccines represent the swiftest developed vaccine in medical history [156, 157]. The initial two received permission from various regulatory bodies for emergency use in China, the USA, and Canada [158]. The FDA approved official marketing for the Pfizer-BioNTech COVID-19 vaccine in August 2021, making it the first vaccine to get such approval. Additionally, on October 29, 2021, it became the first vaccine authorized for children under the age of 11 years. Kitano et al. [159] did a risk assessment of the mRNA vaccine for COVID-19 in children from 6 months to 4 years and found that the benefits of the mRNA vaccine far outweigh any negative effects. The Pfizer-BioNTech vaccine has a dose of 30 μg and is 95% effective, and the Moderna vaccine is 94.5% effective with a dose of 50 μg [160]. After inoculation with homologous SARS-CoV mRNA vaccines, individuals have a superior antibody response than other vaccines [161], and spike antibody assay of the antibody is preferred over nucleoside antibody [162].
Because of the intricate relationships between SARS-CoV-2, its variants, and societal dynamics, environmental factors are crucial to the spread of COVID-19 and make predictive models more difficult to understand [163, 164]. While higher COVID-19 vaccination rates are essential, they alone are not sufficient to significantly reduce mortality; additional health policies and interventions are also necessary [165]. Higher healthcare spending and better medical infrastructure are linked to lower COVID-19 fatality rates, with Western European countries showing stronger outcomes compared to Eastern Europe due to greater per capita health investment [166]. Organizations like WHO, Coalition for Epidemic Preparedness Innovations (CEPI), Global Alliance for Vaccines and Immunization, and International Vaccine Institute (IVI) are crucial in pandemic preparedness, working together to enhance vaccine access and strengthen global responses to infectious diseases [167].
The 2019 pandemic has catalyzed the growth of platforms for mRNA vaccines for the treatment and prevention of several contagious infections, leading to the progressive introduction of a new generation of vaccinations for the general public, which has garnered amplified attention. The vaccines based on mRNA, in contrast to conventional vaccinations, enable the modification of the design of antigens and provide the integration of sequences from many variations to address novel changes in the viral genome. In the future, the mRNA-based vaccination will facilitate the management and prevention of viral diseases and the treatment of other conditions. The foremost vaccination approved by the FDA for COVID-19 was an mRNA-based vaccine because of its benefits, rapid development cycle, absence of cell culture requirements, and excellent immunogenicity [168]. Even machine learning can be applied to data like the number of infections, the turnout in the participation in the vaccination campaign, the respective vaccination plan, and the doses administered to reduce side effects and fatality rate [169]. The COVID-19 pandemic demonstrated that urgent health crises catalyze problem-driven innovation, resulting in rapid breakthroughs such as mRNA vaccines. Addressing these challenges not only advances industrial competitiveness but also contributes significantly to humanity’s ability to overcome pandemic situations [170, 171]. Table 2 lists a few examples of mRNA-based vaccines for viral diseases under clinical trials [172].
List of mRNA vaccines for viral diseases under clinical trials
| Viral disease | NCT identifier | Phase/Status | Delivery route/Formulation |
|---|---|---|---|
| Influenza | NCT05227001 | I/Completed | i.m. injection/NA |
| NCT05426174 | I/Completed | i.m. injection/NA | |
| NCT05052697 | I/II/Completed | i.m. injection/NA | |
| NCT04956575 | I/II/Completed | i.m. injection/LNP | |
| NCT05415462 | III/Completed | i.m. injection/LNP | |
| NCT05333289 | I/II/Completed | i.m. injection/LNP | |
| NCT06744205 | I/II/Recruiting | i.m. injection/NA | |
| NCT06727058 | I/II/Recruiting | i.m. injection/NA | |
| Zika virus | NCT03014089 | I/Completed | Injection/NA |
| NCT04064905 | I/Completed | Injection/NA | |
| NCT04917861 | II/Completed | Injection/NA | |
| HIV/AIDS | NCT00833781 | I/II/Completed | i.d. injection/Dendritic cell |
| NCT02888756 | II/Terminated | Intranasal/Dendritic cell | |
| NCT02413645 | I/Completed | Inguinal intranodal injection/Dendritic cell | |
| NCT06694753 | I/Not yet recruiting | i.m. injection/LNP | |
| NCT05903339 | I/Active, not recruiting | Injection/Ferritin NP and LNP | |
| Rabies | NCT03713086 | I/Completed | i.m. injection/NA |
| NCT02241135 | I/Completed | Injection/RNActive® | |
| COVID-19 | NCT04449276 | I/Completed | i.m. injection/NA |
| NCT05972993 | I/Not yet recruiting | i.m. injection/NA | |
| NCT05144139 | I/II/Completed | i.m. injection/NA | |
| NCT05212610 | IV/Recruiting | i.m. injection/NA | |
| NCT04368728 | II/III/Completed | i.m. injection/NA | |
| NCT04380701 | I/II/Completed | i.m. injection/NA |
AIDS: acquired immunodeficiency syndrome; HIV: human immunodeficiency virus; i.d.: intradermal; i.m.: intramuscular; LNP: lipid nanoparticle; NCT: National Clinical Trial; mRNA: messenger RNA; NA: not applicable; NP: nanoparticle
The mRNA vaccines have emerged as a transformative instrument in combating infectious illnesses, notably exemplified by the efficacy of COVID-19 vaccines such as Pfizer-BioNTech and Moderna. Nonetheless, despite their remarkable efficiency, mRNA vaccination technology has several limitations and solutions.
The fundamental constraint of mRNA vaccines is their volatility at higher temperatures. mRNA is a delicate molecule that rapidly degrades when subjected to heat or inadequate storage conditions. This has created considerable logistical difficulties, particularly for the extensive delivery of the vaccine to isolated or under-resourced regions. The COVID-19 vaccine developed by Pfizer necessitates storage at ultra-low temperatures (about –70°C), demanding specialized freezers and complicating worldwide distribution efforts [173]. Innovative formulations and methods are being created to increase the stability of mRNA vaccines and to proliferate their accessibility in diverse situations.
Since mRNA vaccines provide a robust immune response in several individuals, the degree of immunity may fluctuate among other demographic groups. Variables like age, sex, genetic predisposition, and pre-existing health issues can affect the vaccine’s efficacy. Older people and immunocompromised persons may demonstrate a diminished immunological response to mRNA vaccines, perhaps necessitating booster doses or alternate immunization approaches [174, 175]. In the case of pregnancy, COVID-19 mRNA vaccination is safe, showing no increased risk of adverse outcomes, and is associated with a reduced risk of stillbirth [176].
Despite the good safety profile demonstrated by mRNA vaccines in extensive clinical studies, apprehensions persist over a few side effects. Certain individuals have encountered adverse responses, including anaphylaxis, to constituents in the vaccine formulation, such as LNPs utilized for mRNA delivery into cells. Additionally, there have been a few instances of myocarditis and pericarditis, especially in younger boys after mRNA immunization [177]. These few side effects need continuous monitoring and may undermine public trust in immunizations [178].
The manufacturing of mRNA vaccines necessitates advanced technology and meticulous accuracy. The manufacturing of mRNA, its encapsulation in LNPs, and the requisite quality control to guarantee batch uniformity can be resource-intensive. This has constrained the capacity to rapidly increase output, particularly during the early phases of the COVID-19 epidemic. Moreover, guaranteeing equal distribution is a hurdle, as affluent countries have greater access to these vaccinations than lower-income nations [61, 174, 179, 180]. The development of various mRNA vaccines for COVID-19 paves the path for the development of optimal vaccines for diseases like Tuberculosis (TB), AIDS, influenza, etc. [181].
The long-term efficacy and length of protection of mRNA vaccines, while offering robust defense against illnesses such as COVID-19, remain subjects of ongoing investigation. Preliminary research indicates that immunity may diminish over time, necessitating booster doses. The introduction of novel viral variations is a difficulty, since vaccinations may not provide sufficient protection against these newer strains, necessitating ongoing modifications to vaccine formulations [176, 182, 183].
The mRNA vaccines, despite their promise, encounter several problems such as stability concerns, variability in immune responses, safety issues, manufacturing complexities, and the necessity for long-term effectiveness evidence. Current research and technical progress seek to overcome these restrictions and broaden the worldwide application of mRNA vaccine technology for illnesses outside COVID-19. Data from various countries show that vaccination rates naturally rise with income, peaking around 70% without coercion. Forcing higher rates through strict mandates may harm democracy and cause greater socioeconomic issues than the pandemic itself [184]. Effective COVID-19 crisis management highlights the need for coordinated multi-level governance, timely information sharing, and rapid vaccine R&D to combat future pandemics. Strong leadership, preparedness, and strategic investments are essential to protect public health and socioeconomic stability [185].
In the field of global vaccination technology, mRNA vaccines have quickly evolved from a theoretical breakthrough to a game-changing instrument. Recent discoveries, especially during the COVID-19 pandemic, have shown the enormous potential of mRNA platforms for both therapeutic and preventive applications, after twenty years of research and development. The synthesis, mode of action, delivery methods, and disease-specific uses of mRNA vaccines for significant viral threats, such as COVID-19, Zika, rabies, influenza, and HIV, were all critically investigated in this study. By doing this, it filled in important gaps in the literature, especially the lack of comparative analysis of delivery methods and implications for particular diseases.
This review explains how molecular design, delivery methods, and immune modulation interact to influence vaccine efficacy, adding to the expanding theoretical framework surrounding mRNA vaccine development. The necessity for quick, scalable, and efficient solutions to pandemic emergencies and resistant viral diseases led to the development of mRNA vaccines, which are highlighted as a problem-driven innovation. The study deepens our knowledge of how nucleotide modifications and nanoparticle-based delivery systems affect mRNA stability, immunogenicity, and expression efficiency by combining findings from preclinical and clinical investigations.
Additionally, this study offers a novel conceptual framework for vaccine development that integrates translational medicine, immunology, and nanotechnology. It suggests that mRNA vaccines are not merely an alternative to traditional platforms but a foundational shift in how we approach global immunization strategies, with implications that extend into oncology and other therapeutic areas.
This review highlights the strategic importance of investing in mRNA infrastructure from a managerial and policy perspective, especially for pandemic preparedness and global health equity. The effectiveness of mRNA vaccines during COVID-19 has shown that they can be developed and implemented quickly, which is essential for halting the spread of newly emerging infections. Funding and regulatory support for cold-chain innovations, scalable manufacturing, and region-specific vaccine platforms that target regional disease burdens should be given top priority by policymakers.
The pharmaceutical industry and health authorities around the globe must also adopt flexible regulatory frameworks that can take into account the modular nature of mRNA platforms and their quicker development schedules. In addition, the integration of mRNA technology into routine vaccination programs and public health policy could revolutionize how we prevent and respond to infectious disease outbreaks, especially in resource-limited settings.
Even though the available data is encouraging, the science behind mRNA vaccines has not yet reached its full potential. Future studies ought to concentrate on:
Recognizing the long-term immune reactions brought on by mRNA vaccines, including any possible impacts on immune memory and innate immunity.
Enhancing formulation and delivery methods, especially those that lessen reliance on the cold chain and increase temperature stability.
Looking into how mRNA vaccines might work in concert with other therapeutic approaches, like checkpoint inhibitors or other immunotherapies, to treat cancer.
Investigating mRNA vaccines that are customized for specific immune profiles or local pathogen strains.
Lastly, more studies on self-amplifying RNA platforms and multi-antigen mRNA constructs could lead to more powerful, affordable, and long-lasting vaccines in the future.
Finally, expanded research on multi-antigen mRNA constructs and self-amplifying RNA platforms may pave the way for next-generation vaccines that are more potent, cost-effective, and durable. As the field matures, collaborative efforts between academia, industry, and government will be crucial to unlocking the full potential of mRNA technologies for global health transformation.
AIDS: acquired immunodeficiency syndrome
DCs: dendritic cells
HIV: human immunodeficiency virus
HPV: human papillomavirus
IAVs: influenza A viruses
IFN: interferon
LNA: locked nucleic acid
LNPs: lipid nanoparticles
MHC: major histocompatibility complex
mRNA: messenger RNA
ORF: open reading frame
PBAEs: poly(beta-amino esters)
pDNA: plasmid DNA
PEG: polyethylene glycol
PEI: polyethylenimine
SARS-CoV-2: severe acute respiratory syndrome coronavirus 2
siRNA: short interfering RNA
ssRNA: single-stranded RNA
UTR: untranslated region
VM and AR: Conceptualization, Data curation, Formal analysis, Writing—original draft, Writing—review & editing. YM: Validation, Visualization, Methodology, Formal analysis, Supervision, Writing—review & editing.
The authors declare that they have no conflicts of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
© The Author(s) 2025.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2025. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Imam Nurjaya ... Moh. Anfasa Giffari Makkaraka
Clett Erridge
Marina Yusoff ... Malik Amonov