 Review
Review

Affiliation:
1Department of Biological Sciences, Faculty of Science, Beirut Arab University, Beirut 1107-2809, Lebanon
Email: jamilchahrour@gmail.com
ORCID: https://orcid.org/0009-0000-4916-8873
Affiliation:
2Department of Food and Technology Studies, Faculty of Agronomy, Lebanese University, Beirut 146404, Lebanon
Affiliation:
3College of Engineering and Technology, American University of the Middle East, Egaila 54200, Kuwait
ORCID: https://orcid.org/0000-0002-2070-5855
Affiliation:
2Department of Food and Technology Studies, Faculty of Agronomy, Lebanese University, Beirut 146404, Lebanon
ORCID: https://orcid.org/0000-0003-4984-8531
Explor Neurosci. 2026;5:1006137 DOI: https://doi.org/10.37349/en.2026.1006137
Received: January 10, 2026 Accepted: April 07, 2026 Published: May 14, 2026
Academic Editor: Marcello Iriti, Milan State University, Italy
The article belongs to the special issue Medicinal Plants and Bioactive Phytochemicals in Neuroprotection (Vol II)
Thymoquinone (TQ), the main bioactive constituent of Nigella sativa, has gained great attention for its neuroprotective properties, especially for Alzheimer’s disease (AD), which is a progressive neurodegenerative disorder with limited therapeutic options. This review provides several experimental evidence on the effects of TQ in AD models. The evidences indicate that TQ reduces the amyloid-β accumulation, reduces the oxidative stress and neuroinflammation, and improves cognitive and behavioral outcomes. Additionally, TQ should be able to promote the neuronal survival and neurogenesis while reducing biological markers that indicate brain damage or neuron loss. Although these findings clearly highlight and show the promising therapeutic potential of the TQ molecule in the AD, it is important to note that further in-depth studies are still needed to fully understand its underlying molecular mechanisms and to determine its clinical relevance in patients.
For many centuries, herbal medicine has played a central role in traditional medical systems for the prevention and treatment of a wide range of health disorders and diseases. Different plant organs, including leaves, stems, roots, flowers, fruits, and seeds, have been used extensively as complementary and alternative therapeutic factors. Medicinal plants exhibit various biological activities such as antiviral, anthelmintic, antibacterial, antiallergic, anticancer, antimalarial, antidiabetic, antioxidant, and anti-inflammatory effects, in addition to notable neuroprotective properties [1, 2].
This therapeutic potential of medicinal plants is largely related to the production of the secondary metabolites. These bioactive compounds are classified according to many factors such as their chemical structure, solubility, and biosynthetic pathways. The major classes include alkaloids, phenolics, flavonoids, tannins, terpenes, saponins, glycosides, and volatile oils, all of which contribute significantly to the pharmacological and therapeutic properties of medicinal plants [3].
Among the medicinal plants with neuroprotective properties, Nigella sativa L., which is a member of the Ranunculaceae family, has attracted considerable scientific attention. This plant is cultivated widely across Eastern Europe, the Middle East, and Western Asia. It’s commonly known as black cumin or black seed, Nigella sativa is referred to in Arabic-speaking regions as “Habba Al-Sauda” or “Habba Al-Barakah” and it is incorporated frequently into the daily diet [4]. It is also known as “Cumin Noir” in French, “Echter Schwarzkümmel” in German, “Kalonji” in Hindi, and “Çörek otu” in Turkish.
Historically, Nigella sativa has been referred to as a “miracle plant”, “universal remedy”, and “plant from heaven” by the old ancient civilizations. Archaeological evidence of its medicinal use dates back to the ancient Egypt, where the black seed was discovered in the tomb of Pharaoh Tutankhamun (1333–1323 BC). Furthermore, its significance in traditional medicine is highlighted by the widely cited statements that are attributed to the Prophet Muhammad (PBUH): “Use the black seed, for indeed it is a remedy for all diseases except death,” showing the long-standing cultural and therapeutic value of this plant [5]. Accumulating evidence supports its neuroprotective potential, especially in the context of neurodegenerative disorders.
Chemically, Nigella sativa seeds contain a complex mixture of bioactive compounds, including essential oils, saponins, alkaloids, carbohydrates, proteins, and fixed oils. From a nutritional perspective, the black seed is rich in vitamins, minerals, carbohydrates, fats, and proteins, which play a major role in its health-promoting properties [6]. Also, the seeds contain esters of structurally unique unsaturated fatty acids that are combined with terpene alcohols, accounting for approximately 7% of their composition.
The essential oil content of the Nigella sativa seeds ranges from an average of 0.5% to a maximum of 1.5%, and it is characterized by a high concentration of thymoquinone (TQ) (30–48%), along with p-cymene (7–15%), t-anethole (1–4%), and the sesquiterpene longifolene (1–8%). TQ molecule and its derivatives, including dithymoquinone, thymohydroquinone, and thymol, are considered the most pharmacologically active constituents. Also, additional terpene compounds such as carvacrol, carvone, limonene, 4-terpineol, and citronellol are present in trace amounts [7, 8].
Notably, several studies have reported that the bioactive compounds of Nigella sativa, especially, exhibit antidepressant and analgesic effects, which further supports its therapeutic potential in neurological and neuropsychiatric disorders [9, 10].
Among the bioactive constituents of the Nigella sativa, TQ is widely recognized as the principal compound that is responsible for many of its neuroprotective properties. TQ is a monoterpene benzoquinone molecule formed during plant secondary metabolism from γ-terpinene. Chemically, it is identified as 2-isopropyl-5-methyl-1,4-benzoquinone, a yellow crystalline compound having a molecular formula of C10H12O2 and a molecular weight of 164.20 g/mol. Regarding to its favorable safety profile and low toxicity, TQ has attracted significant interest as a promising pharmacological candidate with significant therapeutic and commercial potential [11].
In addition to its neuroactive properties, extensive experimental evidence has shown that the TQ molecule exhibits a wide spectrum of biological activities, including antioxidant, hepatoprotective, immunomodulatory, antimicrobial, antifungal, gastroprotective, cardioprotective, hypolipidemic, and hypoglycemic effects [12]. It has been reported that TQ protects tissues against oxidative damage induced by free radical-generating agents, such as carbon tetrachloride-mediated hepatotoxicity [13]. Interestingly, TQ shows a concentration-dependent behavior, it acts as an antioxidant at low doses while exhibiting pro-oxidant effects at higher concentrations [14]. Its antioxidant activity is primarily related to its ability to scavenge the reactive oxygen species (ROS), such as superoxide anions, and to enhance the expression of endogenous antioxidant defense enzymes, including superoxide dismutase (SOD), catalase (CAT), and glutathione (GSH) peroxidase [15]. Moreover, these protective effects occur with minimal cytotoxicity toward the normal cells, which supports its therapeutic properties in the neurodegenerative conditions such as Parkinson’s and Alzheimer’s disease (AD) [16].
Considering this wide range of the biological and neuroprotective effects of the TQ molecule, increasing attention has been directed in order to understand its molecular mechanisms that is related to its pharmacological activity. These mechanisms are primarily linked to the regulation of oxidative stress, inflammatory mediators, and key signaling pathways that are involved in cell survival and disease progression.
ROS and reactive nitrogen species (RNS) are highly reactive molecules that are capable of inducing cellular damage through lipid peroxidation, protein oxidation, and DNA modification. Oxidative stress occurs when there is an imbalance between the oxidant generation and the antioxidant defense systems, thus leading to dysfunction of biological systems [17]. TQ has been shown to regulate cellular redox homeostasis in a concentration and cell type dependent manner.
In BEAS-2B cells exposed to cigarette smoke extracts, the treatment with TQ at concentrations of 20 and 50 µM significantly increased the activities of antioxidant enzymes, including SOD, CAT, and GSH reductase (GR), in parallel with elevated intracellular GSH levels [18]. In contrast, in cancer cells, TQ molecule exerts a pro-oxidant effects that contribute to its anticancer activity. For example, in A549 lung cancer cells, the exposure to TQ (20–60 µM) resulted in an increased total oxidant status (TOS) and decreased total antioxidant capacity (TAC), along with the reduced activities of antioxidant enzymes such as SOD, CAT, and GSH peroxidase, therefore promoting oxidative stress selectively in malignant cells [19].
In vivo studies further support the antioxidant potential of TQ. In a bleomycin-induced pulmonary fibrosis rat model, oral administration of TQ (10 and 20 mg/kg for 28 days) significantly decreased the oxidative and inflammatory damage by increasing GSH and SOD levels while reducing malondialdehyde (MDA) concentrations. These effects were mediated through activation of the nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling pathway and suppression of the nuclear factor-kappa B (NF-κB) expression [20]. Additionally, TQ has been reported to inhibit cyclooxygenase-2 (COX-2) expression and prostaglandin production in models of allergic airway inflammation, further supporting its role in the redox and inflammatory regulation [21].
The molecular mechanism of TQ for protecting cells against oxidative damage is summarized in Figure 1.
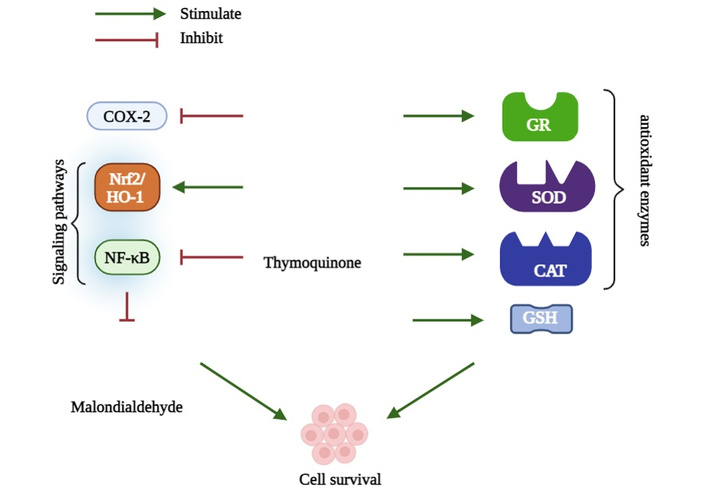
Thymoquinone stimulates antioxidant enzymes, including superoxide dismutase (SOD), catalase (CAT), and glutathione reductase (GR), and activates the nuclear factor erythroid 2-related factor 2 (Nrf2)/heme oxygenase-1 (HO-1) signaling pathway. In contrast, it downregulates cyclooxygenase-2 (COX-2) and nuclear factor-kappa B (NF-κB) expression, which collectively contribute to promoting cell survival. GSH: glutathione.
HO-1 is a cytoprotective enzyme that is involved in heme degradation and plays a key role in protecting tissues against oxidative stress, inflammation, and carcinogenesis. The expression of HO-1 is mainly regulated by the redox-sensitive transcription factor Nrf2, which binds to antioxidant response element (ARE) in the promoter regions of target genes. In normal conditions, Nrf2 is sequestered in the cytoplasm by Kelch-like ECH-associated protein 1 (Keap1), leading to its proteasomal degradation. Upon oxidative stress, the Nrf2 dissociates from Keap1, translocates to the nucleus, and initiates transcription of antioxidant genes.
Experimental evidence shows that the TQ molecule activates the Nrf2/HO-1 pathway. Cells treated with TQ (1–20 µM) for six hours showed significant induction of HO-1 expression at both mRNA and protein levels through Nrf2 activation [22, 23]. Moreover, TQ induced Nrf2 signaling was associated with downregulation of pro-inflammatory cytokines such as interleukin-1β (IL-1β), showing its dual role as antioxidant and anti-inflammatory [24].
The molecular mechanism of TQ for the regulation of Nrf2/HO-1 signaling pathway is summarized in Figure 2.
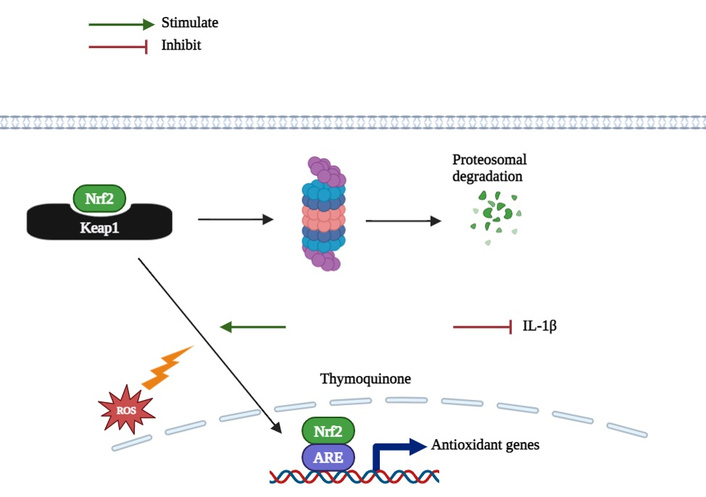
Under normal conditions, nuclear factor erythroid 2-related factor 2 (Nrf2) is sequestered in the cytoplasm by Kelch-like ECH-associated protein 1 (Keap1), leading to its proteasomal degradation. Upon oxidative stress, Nrf2 dissociates from Keap1, translocates to the nucleus, and initiates transcription of antioxidant genes. Thymoquinone activates the Nrf2/heme oxygenase-1 (HO-1) pathway, promotes Nrf2 nuclear translocation, upregulates antioxidant gene expression, and downregulates pro-inflammatory cytokines, such as interleukin-1β (IL-1β), contributing to cellular protection.
TQ exhibits a major anti-inflammatory activity by regulating multiple inflammatory mediators and signaling pathways. Several cytokine profiling studies demonstrated that TQ suppresses the tumor necrosis factor-α (TNF-α) as well as other pro-inflammatory cytokines. In TNF-α-stimulated synovial fibroblasts, TQ treatment (0–1 µM) significantly reduced nitric oxide (NO), IL-6, IL-8, and prostaglandin E2 (PGE2) production in a concentration-dependent manner. These effects were associated with inhibition of Akt phosphorylation, indicating its crosstalk with other intracellular inflammatory signaling pathways [25].
The in vivo studies further confirmed the protective role of TQ against inflammation induced tissue damage. For example, in cisplatin-induced testicular toxicity, TQ decreased the tissue injury by regulating the TNF-α/OTULIN/NF-κB signaling pathway [26]. In a lipopolysaccharide (LPS)-induced neuroinflammation rat model, by using the Evans blue dye method, TQ significantly reduced the blood brain barrier permeability and improved the behavioral performance. These findings indicate that TQ preserves blood brain barrier integrity and reduces inflammation associated with behavioral impairments [27].
TQ has demonstrated marked anticancer activity through regulation of apoptosis, cell survival pathways, and cytoskeletal properties. For example, in breast cancer cells, TQ upregulated peroxisome proliferator-activated receptor γ (PPAR-γ) while downregulating anti-apoptotic genes such as Bcl-2 and Bcl-xL, thus inducing programmed cell death. Molecular docking studies have revealed the interactions between TQ and both polar and non-polar residues within the PPAR-γ binding site, supporting its direct modulatory role [15].
Lipid peroxidation products can form adducts with proteins and DNA, contributing to the progression of tumor. The inhibitory effect of TQ on lipid peroxidation has therefore been proposed as one mechanism underlying its cytoprotective and anticancer effects [21]. Moreover, TQ had inhibited the migration of metastatic human (A375) and murine (B16F10) melanoma cells by suppressing NLRP3 inflammasome activation and reducing caspase-1 cleavage [28].
At the cytoskeletal level, TQ targeted α/β-tubulin proteins in cancer cells, inducing their degradation through the upregulation of the tumor suppressor p73 and activation of caspase dependent apoptosis. Importantly, there were no significant effects observed in normal human fibroblasts. However, in glioblastoma cells, TQ induced apoptosis while inhibiting autophagy, thereby enhancing the anticancer efficacy [28]. Furthermore, in acute lymphoblastic leukemia (CEMss) cells, Treatment of CEMss cells with TQ exhibited strong antiproliferative activity (IC50 = 1.5 µg/mL) by downregulating Bcl-2 and upregulating Bax, leading to apoptosis and inhibition of growth [29].
The molecular mechanism of TQ in controlling cellular apoptosis for normal cells is summarized in Figure 3.
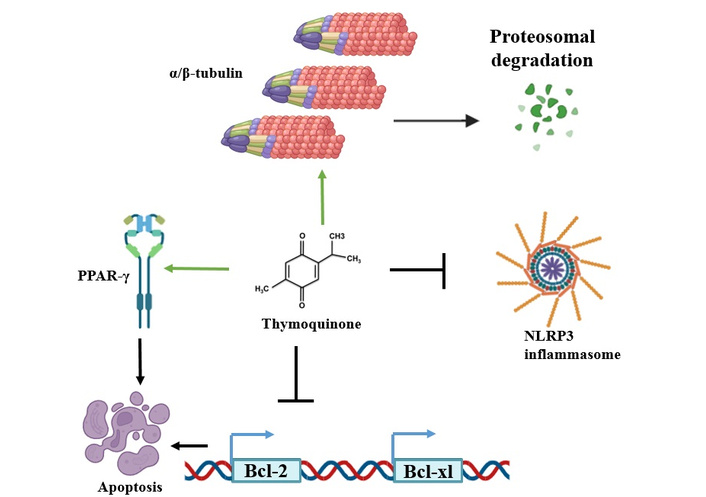
Thymoquinone upregulates peroxisome proliferator-activated receptor γ (PPAR-γ) receptor, that play role in cellular apoptosis and stimulates α/β-tubulin proteins degradation, while suppressing NLRP3 inflammasome and Bcl-2 and Bcl-xL activation. Therefore, promoting cellular apoptosis.
TQ has also demonstrated a notable antiviral activity in both in vivo and in vitro models. In a murine cytomegalovirus infection model, TQ significantly reduced viral loads in the liver and spleen, which is accompanied by enhanced interferon-γ production and increased CD4+ T-cell responses [30]. In vitro studies have reported a significant antiviral activity of TQ against a SARS-CoV-2 strain isolated from Egyptian patients [31]. Also, TQ was also shown to inhibit Epstein-Barr virus replication in infected B cells and reduce the replication of certain coronaviruses [32, 33].
In addition, TQ exhibits broad antimicrobial activity against several pathogenic bacteria, including Streptococcus constellatus, Streptococcus mutans, Gemella haemolysans, and Streptococcus mitis, with inhibition of bacterial biofilm formation. Antibacterial effects have also been observed against Staphylococcus aureus, Salmonella enteritidis, Salmonella typhimurium, Shigella flexneri, Pseudomonas aeruginosa, and Escherichia coli [15]. TQ has further been proposed as a potential agent for controlling the spore forming bacteria such as Alicyclobacillus acidoterrestris.
Regarding the antifungal and the antiparasitic activities, TQ demonstrated potent antifungal effects against eight dermatophyte species, showing greater efficacy than griseofulvin. Additionally, TQ exhibited strong antiparasitic activity against Giardia lamblia and Entamoeba histolytica [34, 35].
The ability of TQ to regulate oxidative stress, inflammation, and cell survival pathways, TQ has been extensively studied for its therapeutic potential in neurodegenerative disorders, especially AD.
AD is a progressive neurodegenerative disorder that is characterized by memory impairment and cognitive decline. Although the precise etiology of the AD remains unclear, but its pathology is mainly defined by two hallmarks: The extracellular neurotoxic senile plaques composed of amyloid-β (Aβ) peptides, which are generated from the proteolytic cleavage of amyloid precursor protein (APP) by β-site APP-cleaving enzyme-1 (BACE-1), and intracellular neurofibrillary tangles that are composed of tau protein filaments. These pathological events initiate a cascade that leads to oxidative stress, synaptic dysfunction, neuroinflammation, and neuronal loss [36].
Several experimental studies highlight the neuroprotective potential of TQ in AD models. For example, TQ has been shown to attenuate Aβ-induced neurotoxicity and to exert anti-inflammatory and antioxidant effects that could mitigate neuroinflammation and cognitive dysfunction in experimental models [37].
Similarly, in a rat model of AD induced by intra-hippocampal infusion of aggregated Aβ (1–42), oral administration of TQ (20 mg/kg/day) promoted functional recovery by enhancing the neurogenesis and decreasing the neuroinflammation through the reduction of interferon-γ (IFN-γ) levels [38].
In addition to its in vivo effects, TQ has demonstrated direct anti-amyloidogenic properties in vitro. Spectrophotometric analysis over a 16-day period revealed that TQ molecule inhibited Aβ (1–40) aggregation in a dose dependent manner, and exhibiting comparable effects to tannic acid, a known anti-aggregation agent. Moreover, TQ had reduced the activation of both caspase-8 and caspase-9, indicating its potential role in suppressing amyloid-induced apoptotic pathways [39].
The neuroprotective efficacy of TQ was further confirmed in the aluminum chloride-induced AD models. Rats receiving TQ concomitantly with AlCl3 showed a significant improvement in spatial working memory, as assessed by the Y-maze test. Also, biochemical analysis revealed significant reductions in serum MDA, Aβ, and tau protein levels, alongside an increase in the GSH peroxidase activity and acetylcholine levels, highlighting the antioxidant and anti-inflammatory mechanisms underlying TQ mediated neuroprotection [40]. Consistently, hippocampal Aβ-injected Wistar rats treated with TQ exhibited improved memory performance in passive avoidance and shuttle box tests, reduced plaque burden in the CA1 region, increased neuronal survival, and prolonged latency times, further supporting the cognitive-enhancing and neurorestorative properties of TQ in the AD pathology [41].
Figure 4 provides a schematic summary of the role of TQ in AD pathogenesis and neuroprotection.
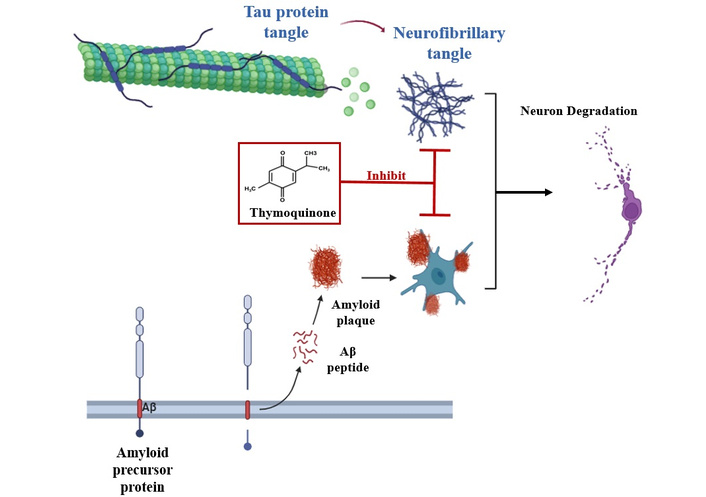
Thymoquinone (TQ) reduces the buildup of amyloid-β (Aβ) peptides and neurofibrillary tangles, which contribute to the pathology of Alzheimer’s disease. By limiting their accumulation, TQ helps prevent the chain of events that cause synaptic problems, inflammation in the brain, and the gradual loss of neurons, supporting overall neuroprotection.
Following ischemic events, neuronal death is primarily driven by the oxygen and glucose deprivation, which triggers oxidative stress, acidosis, and glutamate-mediated cytotoxicity [42]. Oxidative stress is considered a central contributor to the neuronal injury in neurodegenerative conditions, making it a key target for neuroprotective interventions. TQ molecule exhibits neuroprotective effects largely by its potent antioxidant activity, reducing both acute and chronic cerebral pathologies by restoring antioxidant enzyme levels and preventing excessive lipid peroxidation [43].
In AD, oxidative stress plays a major role in the disease progression. Experimental models of transient cerebral ischemia have shown that the daily administration of TQ enhances neuronal survival in the hippocampal CA1 region, which preserves normal levels of GSH, SOD, and CAT, and reduces MDA concentrations in the brain tissues. These protective effects are mediated by TQ-induced activation of the Nrf2 and HO-1 pathway [41].
Further animal studies highlight that TQ has potential against the chemically induced neurotoxicity. For example, intraperitoneal administration of aluminum chloride (AlCl3, 10 mg/kg for 6 weeks) caused impaired motor coordination, anxiety, depression, increased MDA levels, and reduced overall antioxidant capacity. But daily treatment with TQ (10 mg/kg, intraperitoneally) effectively reversed these effects [44]. Additionally, in a scopolamine-induced AD model, rats treated with TQ (20 mg/kg, intraperitoneally, 1 h before scopolamine 1 mg/kg) showed reduction in the brain lipid peroxidation, which supports the role of TQ as a potential adjunctive therapy for AD [45].
Neuroinflammation is the central driver of neurodegenerative processes, which include AD. The activation of microglia is a key event in initiating and maintaining such an inflammatory response, triggered by several stimulations such as infections or traumatic brain injury (TBI). The activated microglia stimulate the NF-κB, which in turn upregulates many proinflammatory mediators, including iNOS, COX-2, and microsomal PGE synthase-1. Therefore, targeting microglial activation, therefore, represents a promising strategy to enhance neuronal survival [46].
Treatment with TQ treatment (2.5, 5, and 10 mM) has been shown to suppress the NF-κB-dependent neuroinflammation in the BV2 microglial cells, which is done by reducing the protein expression of iNOS, as well as inhibiting phosphorylation of IκB, and preventing NF-κB from binding to DNA. However, TQ promoted the accumulation of Nrf2 and enhanced its binding to the ARE, which results in an increased ARE-driven transcriptional activity. These findings indicate that TQ exerts anti-inflammatory effects by the activation of the Nrf2/ARE pathway, which in turn inhibits the NF-κB-mediated neuroinflammation. Moreover, TQ reduced the LPS-induced neuroinflammatory responses by regulating the NF-κB signaling while simultaneously upregulate the activity of Nrf2 [37].
Additional studies have demonstrated that the administration of TQ (2.5, 5, and 10 mM) reduces the production of several inflammatory mediators, including NO, PGE2, TNF-α, and IL-1β in BV2 microglial cells. Automatically, this anti-inflammatory action involves the inhibition of the PI3K/Akt/NF-κB signaling pathway in LPS-stimulated microglia, showing TQ as an important neuroprotective and anti-inflammatory agent in the context of AD [37].
TQ molecule exhibits a pro-apoptotic and cell cycle regulating effects that might have many implications in neurodegenerative conditions, including AD. Cancer preclinical research highlights the TQ’s ability to overcome chemotherapy resistance and enhance other standard therapies. This is done by the induction of cellular apoptosis, regulation of autophagy, endoplasmic reticulum (ER) stress modulation, unfolded protein response (UPR) modulation, and epigenetic regulation [47].
Primarily, TQ molecule suppresses cell proliferation through cell cycle arrest, microtubule organization disruption, and downregulation of pro-survival proteins [48]. The pro-apoptotic action involves mitochondrial dysfunction and activation of caspases, while regulating many pathways such as epithelial-mesenchymal transition (EMT) and oxidative stress, which may also play roles in neuronal protection and survival [49].
At the molecular level, TQ molecule induces cell cycle arrest by inhibiting the cyclin E and cyclin D activation while upregulating other cell cycle inhibitors such as p21 and p27, leading to G1-phase arrest. Notably, the concentration of TQ molecule determines the specific phase that is affected. The higher concentrations can induce G2-phase arrest, while lower doses may cause S-phase arrest in the proliferative cells [48]. Also, TQ upregulates the p53 protein in a time-dependent manner, since it can promote apoptosis via increased Bax expression, and downregulates the anti-apoptotic protein Bcl-2 [47].
These pathways suggest that TQ also may balance or neutralize the aberrant cell survival mechanisms, reduce pathological cellular stress, and promote programmed cell death of dysfunctional neurons, making it a promising molecule for neuroprotection in AD.
It has been revealed that TQ provides antioxidant, anti-inflammatory, and anti-amyloid properties, allowing it to have a major neuroprotective effect in AD and emerge as a promising and potential therapy that can contribute to controlling the disease, so that it could potentially slow down the progression of the disease and improve cognitive function in patients, based on evidence provided in this review.
However, the precise mechanisms of TQ molecule and the interactions in which it contributes to other cellular pathways remain incompletely understood. This includes the possible cross-talk with other pathways that are involved in apoptosis, synaptic plasticity, and neuroinflammation, which may significantly influence the outcomes of the disease.
Moreover, these promising preclinical findings need to be translated into clinical practice, which faces several challenges, such as determining the optimal doses to be used, enhancing the bioavailability of the molecule, ensuring that it’s safe to be used for the long term, and understanding the response of the molecule in different patients, which are considered critical for effective therapy design.
Therefore, future research should focus on uncovering detailed molecular targets of TQ, well-designed in vivo experiments with standard concentrations, and controlled clinical trials to identify its therapeutic efficacy in diverse patient populations. Additionally, future studies should also focus on exploring the combination therapies with other herbal molecules or existing drugs that might provide synergistic effects and improve treatment outcomes for AD.
AD: Alzheimer’s disease
APP: amyloid precursor protein
ARE: antioxidant response element
Aβ: amyloid-β
CAT: catalase
CEMss: acute lymphoblastic leukemia
COX-2: cyclooxygenase-2
GSH: glutathione
HO-1: heme oxygenase-1
IL-1β: interleukin-1β
Keap1: Kelch-like ECH-associated protein 1
LPS: lipopolysaccharide
MDA: malondialdehyde
NF-κB: nuclear factor-kappa B
NO: nitric oxide
Nrf2: nuclear factor erythroid 2-related factor 2
PGE2: prostaglandin E2
PPAR-γ: peroxisome proliferator-activated receptor γ
ROS: reactive oxygen species
SOD: superoxide dismutase
TNF-α: tumor necrosis factor-α
TQ: thymoquinone
The authors would like to state that during the preparation of this manuscript/study, the authors used ChatGPT (OpenAI, version GPT-5) for the purposes of English language editing and grammar refinement with an aimto enhance clarity and readability. OpenAI was not used for generating ideas, analysis, or discussion sections. The authors have reviewed the entire content of the paper and have adjusted and edited all sections and take full responsibility for the content of this publication.
JAC: Writing—original draft, Visualization. MR, ZAB, and AH: Writing—review & editing, Supervision. All authors read and approved the submitted version.
The authors declare no conflicts of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
This research received no external funding.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 1304
Download: 31
Times Cited: 0
