Review
Review

Affiliation:
1Department of Biotechnology, Thapar Institute of Engineering and Technology, Patiala 147004, Punjab, India
Email: spal_phd25@thapar.edu
ORCID: https://orcid.org/0009-0000-2483-9597
Affiliation:
2Department of Microbiology, Sentiss Pharma Private Limited, Nalagarh 174101, Solan, Himachal Pradesh, India
ORCID: https://orcid.org/0009-0004-6173-0995
Affiliation:
3Department of Bio-Sciences and Technology, MMEC, Maharishi Markandeshwar (Deemed to be University), Mullana, Ambala 133207, Haryana, India
ORCID: https://orcid.org/0000-0002-9257-927X
Explor Drug Sci. 2026;4:1008166 DOI: https://doi.org/10.37349/eds.2026.1008166
Received: March 27, 2026 Accepted: May 17, 2026 Published: July 07, 2026
Academic Editor: Ralf Weiskirchen, RWTH University Hospital Aachen, Germany
Today, researchers have already made great progress in finding drug-based clinical solutions against microbial infections. It has become an essential part of a healthy human lifestyle. High antibiotic consumption has accelerated antibiotic resistance in microbial species (multi-drug-resistant microbial strains, like Staphylococcus aureus and Mycobacterium tuberculosis, have already been reported). Loss of microflora is also associated with the heavy and unnecessary use of drugs. This review presents bacteriophage as an alternative to antibiotics. The supporting bacteriophage characteristics include bacteriophage lytic mode of replication, specificity towards its host, bacteriophage mass production, and bacteriophage genetic modification (BRED and CRISPR) to make it capable of degrading microbial biofilm. The author has also tried to inculcate previous work that has already been done with bacteriophages for some clinical therapies. Potential administration routes (oral, intravenous, and intraoperative) used in clinical therapies are discussed.
We are quite aware of the normal flora inhabiting human beings along with some of their advantages, like the secretion of important gut enzymes for food digestion [1] and immune system training. They have complex alternative metabolisms to inhabit changing habitats. Some bacterial fermented products, such as short-chain fatty acids, act like immunomodulators for tissue-specific macrophages in the gut. This interaction of macrophages with short-chain fatty acids inhibits the mechanistic target of rapamycin (mTOR) and increases antibacterial activity [2]. But we also have some diseases caused by the excess growth (infection) of pathogenic bacteria or opportunistic growth of commensals, their interference with the body’s normal physiological activities, toxin production, consumption of essential nutrients and making us deficient for them, evasion of the immune system (Mycobacterium tuberculosis evasion in macrophages and granuloma formation) [3], invasion of the first line of defence or physical barriers like skin, direct tissue degradation, or causing inflammatory reactions [1].
Generally, antimicrobial drugs are used in the healthcare sector to treat diseases caused by microbial infections. Drugs are chemical substances mostly obtained from microbes, plants, marine organisms [4], or synthesized by a series of reactions. Microbes produce drugs to gain a competitive advantage over other microbes and to preserve resources. After Sir Alexander Fleming discovered the first antibiotic (penicillin), organisations began producing it in bio-fermenters using the fungus Penicillium chrysogenum [5]. Penicillin belongs to the family of beta-lactam and acts specifically on the cell wall of bacteria, interfering with its biosynthesis by inhibiting transpeptidase (responsible for catalysing the bacterial peptidoglycan cross-linkages) [6]. The drug is not a primary metabolite, secreted outside the cell (secretory in nature), and is not a must for the survival of the producing microbe. The commercial usage of drugs against microbial infections is a revolution in the healthcare sector. Drug usage impacted life expectancy significantly in the United States of America (USA) by changing the average life span of humans from 47.3 years to 78.93 years from 1900 to 2020 [7]. In the period between 1920 and 1946, soon after the discovery of penicillin, 14 percent of Staphylococcus isolated from patients were found to be resistant [8]. Commercial production of drugs and their poorly controlled usage increase microbial interactions with drugs as compared to the interaction that occurs naturally [9]. As a result, microbes get a chance to evolve faster in the selective pressure. Antimicrobial resistance (AMR) is a natural phenomenon that occurs in the environment. Microbes adapt to changes against the antimicrobial drugs for survival. These changes include gene mutation or genetic transfer among bacterial cells via horizontal gene transfer (for example, Staphylococcus aureus is getting resistant to methicillin) [1, 10–12], expression level of specific proteins to avoid drug intake, altering drug binding receptors, and changes in the membrane permeability. Bacterial metabolism is also a crucial factor in its sensitivity or resistance to antibiotic drugs [12, 13].
To check the bioactivity and effectiveness of drugs, various assays like high-performance liquid chromatography (HPLC), fluorescence immunoassay, radioimmunoassay, and microbial assays (disc-diffusion method) are employed in the pharmaceutical industry. The minimum concentration of an antibiotic that can effectively inhibit half of the targeted microbial population is called the minimum inhibitory concentration (MIC), and every pharmaceutical industry is working on this key principle for manufacturing drugs. The drug usage must be prescribed accordingly [14].
Reasons for developing antimicrobial drug-resistant strains include excess usage of drugs in acute infections, non-compliant disposal, making it available for the microbes while working in the laboratories, and lack of awareness (drug usage without prescription even in case of acute viral infections). In some cases, it was noted that doctors prescribed drugs without serious need. Literature has mentioned the need for proper professional training for the prescriber and appropriate educational awareness in the people to avoid antibiotic usage unless necessary [15]. Completion of the antibiotic regimen is also crucial to prevent the antibiotic movement into the surrounding environment and prevent AMR. Researchers investigated that after administration of 400 mg of fleroxacin and 800 mg of pefloxacin to a healthy person. Antibiotics can be recovered from their urine and it can still create a selective pressure in the surrounding. Significant bactericidal activity of recovered antibiotics against Escherichia coli ATCC 25922, Klebsiella pneumoniae, E. coli, Proteus mirabilis, Pseudomonas aeruginosa I, P. aeruginosa II, Enterococcus faecalis, Staphylococcus saprophyticus, S. aureus, Streptococcus group B sp. has been reported [16]. Biotic and abiotic agents detoxify drugs in the biosphere, like antibiotic conversion to inorganic minerals by structural alteration in the presence of environmental factors (pH, temperature, and light). For example, the beta-lactam bond degrades in a pH-dependent fashion in aqueous solution, and its chemical structure also plays a role [17, 18]. Hydrolysis of clavulanate and carbapenems is relatively easy due to their Woodward-Fieser value (a value of chemical reactivity) [18, 19]. Antibiotic detoxification and degradation are reported to be mediated through oxidation in the wet clay, and their rate is found to be increased by pH and light. Ions such as hydroxyl radicals, Cu+, and Cu2+ are also involved [18, 20, 21]. The human liver also works in drug detoxification, converting toxic drugs to less toxic (water-soluble) compounds that are further excreted [22]. Viruses are the most numerous (up to 2.5 × 108 phage/mL of natural water) [23, 24], acellular particles of protein coat containing their genetic material. The International Committee on Taxonomy of Viruses deals with the classification and nomenclature of viruses. Virus classification is mainly based on the genetic material, host type, and capsid structure. They are obligate parasites [25, 26]. Due to the lack of conserved sequences in the viral genome, the exact phylogenetic relationship with other domains of life is complicated to analyse [27].
Viruses that use bacteria as their host are known as bacteriophages or phage particles [28–30]. The typical structure of the bacteriophage includes nucleic acid covered by a protein capsid. Bacteriophage-like Caudoviricetes have legs, a neck, and tail-like appendages [31]. Bacteriophage shows two types of life cycles. The lytic cycle constitutes attachment of the phage to bacteria and introduction of its genetic material into the bacterial cytoplasm [8]. Followed by the hijacking of bacterial machinery for new phage assembly. A new particle lyses the bacterial cell and gets out of it to search for a new bacterial host. On the other side, the lysogenic phage genome gets integrated into the bacterial genome and starts replicating with it for several generations, and then they induce bacterial lysis again [32, 33].
Bacteriophage can be used as a biomarker for the detection of specific bacterial species [34] and to facilitate horizontal gene transfer in specific bacterial species via transduction [35]. Phage particles are used to target pathogenic bacterial cells in the human body and the food industry [36]. Food and Drug Administration (FDA) and the United States Department of Agriculture (USDA) have approved bacteriophage application in the food industries mainly for Salmonella, E. coli O157:H7, and Listeria monocytogenes [37]. Key differences between antibiotics and bacteriophages are mentioned (Table 1). This shows bacteriophage's advantage over antibiotic treatment.
Key differences show bacteriophages as a better option for the clinical treatment of bacterial infections.
| Antibiotics | Bacteriophages | References |
|---|---|---|
| They are specific for their site of action. But not for the targeted host. For example, neomycin restricts protein synthesis mainly in Gram-negative bacteria, but it is not sensitive to specific bacterial species. | Have highly specific surface proteins that can recognize their host at the species and strain level (bacterial species) through receptors present on host outer structures, including peptidoglycan, teichoic acid, glycolipids, and capsule. | [38–42] |
| Antibiotics generally have a broad range of action. It also affects other bacterial commensals badly, causing microbial imbalance and increasing the chance of secondary infection. | A narrow range of bacteriophage host selection ensures very low effects on normal flora and no selective pressure. Therefore, endogenous bacteria remain protected and prevent multidrug resistance. | [43–46] |
| An antibiotic is a chemical (drug) extracted from an organic source (mainly microbes) or synthesised by chemical reactions. | Bacteriophages can be used as cargo for transferring antibiotic genes inside the bacterium to bypass the bacterial membrane-alteration mechanism to become resistant. | [47] |
| Antibiotics are toxic to consume. Their side effects and complications with longer usage include intestinal disorders, allergies, and secondary infections. | Bacteriophage genes causing toxicity can be altered or removed using genetic engineering. | [47, 48] |
| Antibiotic prescription considers the minimum inhibitory concentration of the infection-causing microbe. Physicians use to prescribe it in a regimen. | Bacteriophage replicates itself inside the host. Its single dose maintains a sufficient number to kill bacterial infection. | [49] |
| Antibiotic discovery and modification are time-consuming processes with a very high cost. | Bacteriophage present in abundance. Their isolation and modification are comparatively less costly. | [50] |
According to the WHO (World Health Organisation), 7 lakh people die per year because of AMR. AMR is a natural characteristic of bacteria. They induce changes in their genome to become resistant to a particular antibiotic present in the surroundings. Excessive use of antibiotics in industries creates selective pressure for resistant bacteria. This results in the transformation of a bacterial pathogen into multidrug-resistant [51, 52]. WHO issued a list of priorities for antimicrobial-resistant pathogens in 2017. This includes Salmonella sp. (fluoroquinolone-resistant), Campylobacter (fluoroquinolone-resistant), S. aureus (methicillin and vancomycin-resistant), Enterococcus faecium (vancomycin-resistant), Neisseria gonorrhoeae (cephalosporin and fluoroquinolone-resistant), M. tuberculosis, P. aeruginosa (carbapenem-resistant), Acinetobacter baumannii (carbapenem-resistant), Helicobacter pylori (clarithromycin-resistant) [53–57]. This has increased the demand for new specialized antibiotics or other antibiotic alternatives. Researchers are now paying attention to non-toxic chemical, physical, and biotic agents to replace antibiotics. In 2017, the Wellcome Trust Commission documented antibiotic alternatives, including bacteriophage, to overcome their side effects [58]. Fredrick Twort first discovered phage particles independently in 1915. Early, they also observed the antibacterial properties of phage. Successful applications of phage in the personalized treatment of A. baumannii, Mycobacterium abscessus, and P. aeruginosa have already been reported [47]. Researchers used bacteriophage in association with antibiotics against the bacterium extraintestinal pathogenic E. coli (ExPEC). This lowers the MIC of antibiotics by enhancing the drug’s bacterial growth inhibitory effect and suppressing the emergence of antibiotic-resistant cells [49].
Bacteriophage tail recognizes its host specifically. Bacteriophage cocktail therapy exploits a mixture of bacteriophages with different specificities of tails to target a wide range of bacteria [45, 59]. Phage SP22 and SP21 have binding capability for the two different outer membrane proteins of enterotoxigenic strain E. coli O157:H7, that is, lipopolysaccharides (LPS) and outer membrane protein C (OmpC) [42]. The range of pathogenic bacterial strains for bacteriophage cocktail therapy also includes E. coli and Staphylococcus to treat ear infections and gastrointestinal infections [60].
Pharmacokinetics states the process of drug elimination from the body. This helped the physician to describe the dosage of the antibiotic in the regimen. Dosage plays a vital role in maintaining the minimum administration of drugs that can efficiently kill the microbial infection. However, a tiny amount of bacteriophage dose is enough in a single administration. Because bacteriophage replicates there until the bacterial population is killed completely [8, 61], also called auto-dosing [62]. A threshold density of ≥ 104/CFU/mL of host bacterium is generally required for the auto-dosing of bacteriophage [63, 64]. Bacteriophage also helps in the clearance of S. aureus infection by mobilising the cells from myeloid and lymphoid lineages, recruiting the immature neutrophils from the bone marrow, and helping them to mature for their function in the peripheral circulatory system. This was demonstrated on an immunosuppressed mouse model by administering phage to it [65].
Some bacteriophage characteristics must be considered that should either be present in the bacteriophage or need to be induced. For example: bacteriophage life cycle (replication within a host bacterium), host specificity, bacteriophage administration, mass production, and biofilm degradation. Bacteriophage and its respective bacterial host are coevolving over a very long time. Therefore, their interactions can’t eliminate either population. Bacteriophage eliminates the bacterial host population through the synergistic effect of the immune system. The immune system crucially ensures the prevention of the emergence of phage-resistant bacteria [66].
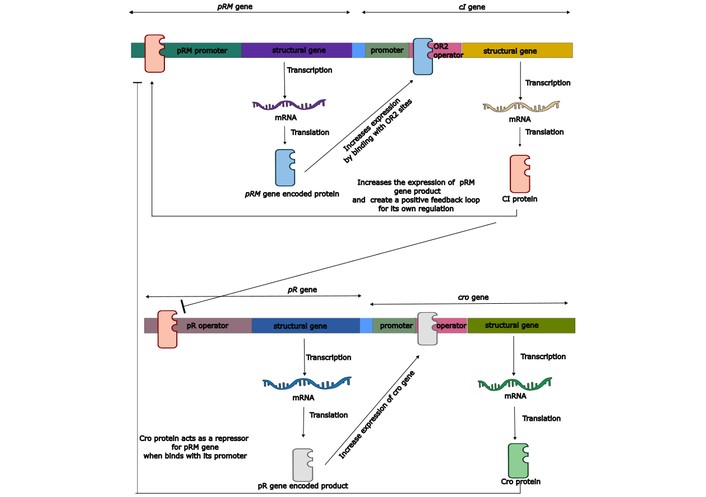
In the lytic cycle, the bacteriophage first inserts its genome into the host bacterial cell. Then, integrate its genetic material into the bacterial chromosomal genome. Integration is followed by the use of bacterial cellular mechanisms by the bacteriophage genome to synthesise numerous copies of its own genetic material and the required proteins for capsomers. Intracellular assembly of bacteriophage particles takes place, followed by their release from the bacterial host cell to infect another one. Bacterial lysis is a result of bacteriophage endolysin breaking the peptidoglycan, followed by its destabilisation and rupture due to osmotic pressure. Bacteriophage assembly is a very complex process, including protein-protein and protein-nucleotide interactions. It is also a model system for the investigation of biological assemblies. The lytic cycle used by the bacteriophage prevents transduction in all circumstances [67–69]. Genetic manipulation of the life cycle of bacteriophages is possible by understanding their genetic regulatory mechanism. Mainly, two bacteriophage genes regulate the phage life cycle. One is the cI gene, which controls the lysogenic life cycle of the phage. The cl gene produces a CI protein through transcription and translation. CI protein acts like a transcriptional repressor. It binds to the two gene operators named pR and pL to inhibit their respective gene expression. The pR promoter gene is responsible for the lytic cycle of bacteriophage by derepressing the cro gene. The second task is to stimulate its expression by interacting with the pRM promoter. The cro gene produces the Cro protein that inhibits the pRM promoter and inhibits the expression of the cI gene [70]. Knocking out the cI gene can be performed to get a phage particle with the characteristic of performing a lytic cycle only. Typical genetic regulation of the bacteriophage replication cycle is well illustrated (Figure 1). BRED (bacteriophage recombineering of electroporated DNA) is an in vivo technique and involves the recombination of very short homologies using the molecular machinery of the bacteriophage itself. First performed in the mycobacteriophage in the year 2008. Che9c, gp60, and gp61 proteins are present in the mycobacteriophage, and it is similar to the RecE/RecT system found in the Rac prophage of E. coli. A substrate (ssDNA) and the prophage DNA template are electroporated into an electrocompetent cell that expresses recombination proteins (gp60 and gp61). Selection can be done with either polymerase chain reaction (PCR) or by plating the competent cell for analysing the plaque. This technique can be used for deletion, knockouts, replacement, and point mutations in the viral genome [71–74]. This technique can be employed for the knockout of repressor genes of the bacteriophage lytic cycle. Genome sequencing and other bioinformatic tools help to search for and identify desirable and undesirable genes. This can be done by homology search of the phage genome with the genes (toxin genes, antibiotic genes, integrase, and repressors associated with the phage lysogeny) existed in the databases [75, 76].
Bacteriophages are very specific for their host cell. Gene editing helps in designing desired bacteria-targeting bacteriophages with selective specificity. Specific bacteriophage tail genes can be expressed in another bacteriophage genome to get the desired host specificity. The end product will be a recombinant bacteriophage that would be able to express the tail structures of interest and can target multiple bacterial species or any specific bacterial species. For example, Bacteriophage T4 (Escherichia coli Virus T4) infects E. coli using tail fibres, and the genes working behind this specificity are gp34 and gp37 (gp stands for gene product). The efficiency of bacteriophage T4 is almost 1.0. Every bacteriophage particle successfully replicates/insert its genetic material after getting adsorbed on the E. coli surface. In the interaction between E. coli and bacteriophage T4, the glucose residue on the core lipopolysaccharide of E. coli serves as a receptor for host recognition [77]. Clustered regularly interspaced short palindromic repeat (CRISPR) was first documented in E. coli in the year 1987 as a defence mechanism against bacteriophage. This can cleave the bacteriophage genome in a sequence-specific manner. Researchers had already used CRISPR as a gene editing tool on Vibrio natriegens TT4P2 in the host bacterium V. natriegens TT4 to achieve gene deletion and replacement. They successfully deleted the orf6 gene of size 292 bp, the orf45 gene of size 162 bp, and replaced the orf6 gene with orf39 [23]. CRISPR is a promising technique to make various recombinant bacteriophages with our desired genetic alterations. This will also counter the act of bacteriophage against human-associated bacterial flora [78, 79]. BRED can also be used for the same. Clinical isolation and identification of the infectious bacteria are crucial for personalised bacteriophage therapy. The phage biobank helps in considering several bacteriophages against the infectious bacterial strain, and high-throughput screening helps in finding specific phage matches with the bacterial host [62].
Biofilm is a fundamental defensive mechanism of bacteria against antibiotics. This prevents antibiotics’ action on bacteria. Biofilms can be observed on places like dental plaques, medical devices, and foodstuffs [80, 81]. It is a slimy layer formed as a result of bacterial extracellular secretions. It mainly comprises exopolysaccharides like N-acetyl-D-glucosamine (dPNAG) exopolysaccharide, alginate exopolysaccharide, psl exopolysaccharide, amylose like glucon. Enzymes like dPNAG glycoside hydrolase (dispersin B), alginate glycoside hydrolase, Psl glycoside hydrolase, and amylose glycoside hydrolase target these exopolysaccharides, respectively [82]. A capsule with the above-integrated enzymes can be developed by using artificial heterodimeric or even monomeric antibodies, which can be fused with the coat protein of a bacteriophage or other capsule-like structures through the constant region of the antibody. This fusion can be achieved by the phage display system [83]. Phage display exploits the screening of a large number of peptides for their binding ability to target molecules. The desired protein (bacterial surface-binding receptor protein or antibody) encoding gene can be inserted between the genes for phage capsids. At the time of phage harvesting, phage particles express the desired protein molecules integrated into their capsid [44]. Use of phage depolymerase is also a good concept for biofilm degradation. Phages infecting mainly E. coli K1, E. coli K20, K. pneumoniae K22, and A. baumannii K26 encode depolymerases [84–86]. The phagemid technique can be utilised for preparing antibodies fused to the coat protein. A phagemid is a genetically engineered DNA molecule with an integrated bacteriophage genome, plasmid replication site, restriction sites, selective markers, and an antibody-encoding gene [87]. The transformation of the phagemid allows it to replicate in a competent bacterial cell to produce a bacteriophage of interest. Not every bacterial cell is competent to receive foreign DNA molecules like plasmids, chromosomal DNA, and free molecules of DNA present in the environment [88]. However, bacteria can be made competent through treatments, such as the calcium chloride method, in which bacterial cells are treated with calcium chloride at low temperatures and suddenly treated with heat (42°C) for 30–60 seconds. The calcium ions neutralise the negative charge present on the bacterial cell membrane and the bacterial DNA. Neutralisation of negative charge and heat shock treatment help bacterial membranes to take up foreign DNA molecules. Almost all divalent cations, such as calcium or magnesium, enhance the transformation. The presence of magnesium ions increases the transformation by 15–20-fold [89, 90]. Electroporation creates temporary pores in the bacterial cell membrane and makes the membrane competent to take foreign DNA molecules [91].
S. aureus, Thermus thermophilus, N. gonorrhoeae, Vibrio cholerae, H. pylori, Haemophilus influenzae, and Bacillus subtilis are naturally competent species for transformation [92]. Prokaryotic antibiotic genes like actinomycin D and mitomycin (which block the prokaryotic translation by interrupting the elongation factor) can be used in the phagemid to kill the bacteria after coming into contact with a phage-infected bacterium [93]. In this way, bacteriophage acts as a specific drug delivery carrier for other bacterial strains present nearby. It restricts its own growth and the growth of surrounding non-infected bacteria through antibiotic production [94]. Researchers enhanced dispersin B digestion by engineering a bacteriophage having a gene of the dispersin B-degrading enzyme. Bacteriophage T7 against biofilm formed by E. coli has been engineered [95]. Schematic diagram showing the bacteriophage covered with a capsule with integrated enzymes (Figure 2). Human innate and acquired immune cells interfere with phage therapy and decrease the effectiveness of the phage on bacteria. In an experiment, the ability of intracellular killing by phagocytes in a patient suffering from chronic bacterial infection is considered. It was found that the intracellular killing ability is higher in patients after phage therapy than before the therapy [96]. This experiment shows that bacteriophage recovers this ability of intracellular killing of pathogens by phagocytic cells. Another experiment resulted in a higher killing effect of an engineered fluorescent K1F-GFP bacteriophage having specificity for E. coli when it entered the T24 human urinary bladder epithelial cell [97]. This showed that bacteriophage can work on bacteria in the presence of the human immune system, and it may even collaborate with the immune system to kill bacteria.
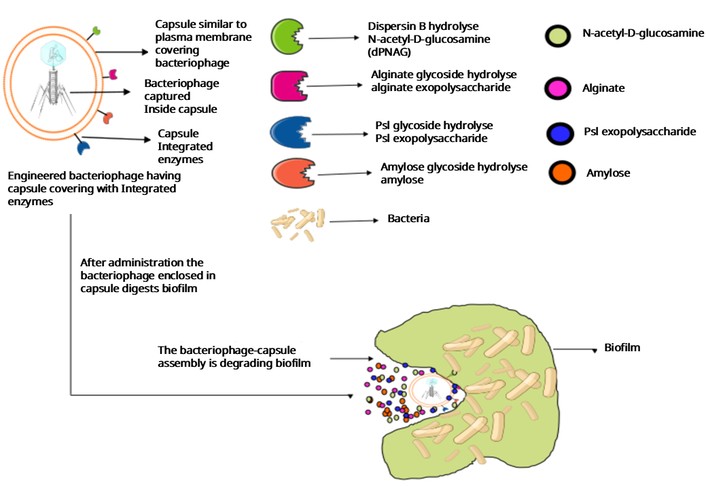
A diagrammatic representation of a hypothetical structure of a capsule containing bacteriophage and integrated enzymes to target biofilm polymers.
Administration is crucial to the efficacy of a drug, vaccine, or any other antimicrobial substance. Different routes, including oral, intravenous, topical, intrarectal, intraoperative, and nebulization, are available for bacteriophage administration [53, 98–100]. The first successful bacteriophage administration was performed in 1921 in France (Hôpital des Enfants-Malades, Paris) against Shiga dysentery in a child. The effectiveness of bacteriophage therapy is highly controversial [101]. Oral bacteriophage administration has poor stability in the acidic environment of the stomach. HCl present in the gastric secretion and digestive enzymes degrade bacteriophage capsomers (protein coat) [11]. Bacteriophage stays in the intestine for a short time, not long enough for effective host attachment. Therefore, repetitive administration is needed for effective phage therapy [102]. The bile salts and carbohydrates present in the intestine block divalent ions, that’s must for bacteriophage replication. Resulting in the reduction of bacteriophage replication in the host bacterium. Unlike drugs, bacteriophages are generally not absorbed by the intestine and cannot enter the blood circulation. This limits oral administration to treat bacterial infections present only in the gut [51, 53]. Bacteriophage encapsulation (using liposomes, electrospun fibre-encapsulation, and polymers) protects phage particles from gastric secretions. Liposome-encapsulated bacteriophages are covered with a positively charged liposome. It protects the bacteriophage from the acidic pH of the human body, acts as a promoter for the mucosal adhesion of phage particles, and increases the retention time of the bacteriophage in the intestine. The flaw of liposome encapsulation is its nature of conglomeration. Liposomes form large conglomerates through adhesion with each other. Conglomeration compromises the stability of the phage particle and causes non-specific interactions with the target host cell. In electrospun fibers-encapsulated bacteriophage therapy, bacteriophages are attached to the surface of electrospun fibers. This is one of the easiest ways to introduce the bacteriophage to the local site of infection. It helps to identify, detect, capture, kill, and immobilise the targeted bacteria. Electrospun-encapsulation also causes the dehydration of the phage particle and leads to its destruction. Microneedle therapy is another form of bacteriophage administration that involves direct penetration of the needle and introduces the bacteriophages to the site of infection. However, this procedure is very difficult for the patient to accept. Polymers-encapsulated bacteriophages are prepared by the chemical modification of biopolymers like polyamines, polyamides, polycarbonates, alginates, and aliphatic polyesters. Polymer increases the acid stability of bacteriophage in the gastric cavity. For example, alginate polymer encapsulation with the addition of guar gum, pectin, and chitosan. Biopolymer-encapsulated phage does not stimulate proinflammatory cytokines and antibody production. But stimulates phagocytes in the blood circulation [51, 103–112].
The intraoperative route involves direct bacteriophage administration at the site of infection. In the year 2012, a 76-year-old patient was suffering from an aortic aneurysm. During the aortic arch replacement surgery, he got an infection of P. aeruginosa. The infection was diagnosed through a deep wound and blood culture. After the recurrence of the same bacterial infection on the same site. He was treated with a mixture of phage OMKO1 and ceftazidime (0.2 g/mL) into the mediastinal fistula. The patient had never faced the recurrence of P. aeruginosa infection again [113]. A 62-year-old male was suffering from fulminant pleural empyema after LVAD (left ventricular assist device) implant. He was diagnosed with the infection of S. aureus through a wound swab. There was no improvement after the antibiotic treatment with daptomycin (500 mg) that was administered intravenously once per day. Finally, 14 doses of Staphylococcus phage CH1 (1 × 109 PFU/mL) were administered via drainage at the site of infection every 12 hours, along with the same antibiotic regimen. After one week of Staphylococcus phage CH1 administration, eradication of S. aureus from the site of infection was confirmed by microbiological assay. Another 66-year-old female patient was suffering from a sternal wall healing disorder. She had aortocoronary bypass and mitral valve replacement surgery. Wound swab confirmed the infection of E. coli, and the patient’s condition was not improving with antibiotics. 600 mg of clindamycin three times per day was administered orally. A mixture of Escherichia phage V18 and Escherichia phage ECD7 was administered intraoperatively, mixed with fibrin glue. E. coli was not detected in further microbiological assays. Similar case of a patient (13-year-old male) who had a double lung transplant and developed sternal wound abscesses. Infection with P. aeruginosa has been confirmed from a wound swab. The patient did not recover after using antibiotic treatment of 187.5 mg avibactam, 750 mg ceftazidime intravenously three times per day, and 2 MIU colistin twice per day. He recovered after the administration of Pseudomonas phage PA5 and Pseudomonas phage PA10 intraoperatively mixed with fibrin glue [114].
The intravenous route of administration involves the introduction of medicine, drug, or food supplement (vitamins and calcium, etc.) directly into the blood vessels. A 77-year-old male patient got infected with multi-drug-resistant A. baumannii (a gram-negative coccobacillus) after the craniectomy. A collection of 104 A. baumannii bacteriophages was analyzed against the isolate of A. baumannii collected from the patient and cultured in the tryptic soy broth by NMRC (Naval Medical Research Center-Frederick). The screening of the isolate and bacteriophage collection was also performed at NMRC. Only 5 phages were found to be active against the A. baumannii. The bacteriophage (2 × 1010 PFU/mL, endotoxin level of 3.5 × 105 endotoxin units/mL) in 4 mL of lactated Ringer’s (intravenous saline) was administered intravenously using a central catheter (inserted peripherally). The frequency of administration was once every two hours for 8 days. There were no further signs of infection, and the craniectomy site healed. Further urine, blood, and respiratory cultures were analysed and found negative for A. baumannii [115].
Bacteriophage can also be administered through oral and topical routes (directly on the surface or in the layer of skin) to make its availability easy, non-invasive, and with high levels of patient compliance [116]. A 16-year-old boy suffering from Netherton syndrome (a rare congenital skin disorder) was treated with an undefined mixture of bacteriophage. He showed a reduction in atopic eczema (a chronic inflammation of the skin characterised by swelling, dryness, itchiness, and redness). Used bacteriophage mixtures contained phage for Proteus, Streptococcus, Staphylococcus, Pseudomonas, Escherichia, and Enterococcus. The patient’s health improved with increased joint mobility and reduced skin irritation [117, 118].
Another challenge is the mass production of bacteriophage. Organisations are continuously trying to produce bacteriophage-based products. AmpliPhi Biosciences Corporation has focused on its product named Phagoburn for the treatment of wounds inhabited by E. coli or P. aeruginosa. This organisation has also been working on bacteriophage-based treatment against skin infection of S. aureus. Countries like Russia, Poland, Belgium, and Georgia have used phage therapy for decades due to its positive results [36, 119]. AgriPhage™ is for agricultural lands against plant infections. LISTEX™ P100 and EcoShield™ are used to prevent the growth of L. monocytogenes and E. coli O157:H7, respectively [120]. Traditionally, shaker flasks or stirred tank bioreactors were used for bacteriophage cultivation. Batch, semi-continuous, and continuous modes of cultivation are now optimised for bacteriophage cultivation in bioreactors [121]. A semi-continuous two-stage self-cycling process has been developed to overcome the disadvantages of the above mode of cultivation. In this process, the host bacteria are used to cultivate in the first bioreactor in batch mode. Just before the stationary phase, half of the media from the first bioreactor was introduced into the second bioreactor for bacteriophage cultivation [122]. Host bacterial growth has a crucial impact on the bacteriophage cultivation. Changing the physiological condition (temperature, pH and substrate concentration for bacterial growth) of host bacteria provides different concentrations of bacteriophage [119, 123]. The generalised procedure for phage cultivation is difficult to develop. The literature mentioned three pipelines of phage production (off-the-shelf, pharmacy compounding, and on-demand de novo). Off-the-shelf pipelines constitute the formulation of fixed phage compositions under good manufacturing practices, either against a single bacterial strain or multiple bacterial strains [124]. When microbial isolates of patients are found to be insensitive to the phage used in the fixed phage products. Then the second pipeline is used by the licensed pharmacist. A physician can manufacture a phage product according to the personalised needs of the patient [125]. In this pipeline, the sensitivity of the phage screened against the infectious isolate of the patient from the phage collection and libraries maintained by the institution (Queen Astrid Military Hospital) [126]. In an on-demand de novo pipeline, the sensitive phage particles against the infectious isolates are isolated from the site of infection itself. This pipeline provides good results and better efficacy [40, 127, 128].
In 1919, a child was treated with phage therapy against bacterial dysentery caused by Shigella in France. Felix d’Herelle isolated a microbe having anti-Shigella activity from the stool and urine sample of a patient who had just recovered from bacillary dysentery. Reinoculation of the isolate into the Shigella culture stopped the growth and lysed Shigella cells. This pattern didn’t repeat when inoculating the isolate with the typhoid and paratyphoid cultures. Even the pattern of this lysis didn’t repeat in the case of heat-killed Shigella. It was concluded that the isolate is an obligate bacteriophage having very high specificity towards Shigella [129, 130]. Various clinical implementations of bacteriophage therapy are depicted in Table 2.
History of bacteriophage therapy studies.
| Year | Country | Pathogen | Disease | Description |
|---|---|---|---|---|
| 1919 | France | Shigella | Bacterial dysentery | Treatment of children suffering from severe dysentery using previously isolated phages. |
| 1921 | France | Staphylococcus | Carbunculosis and furunculosis | The injection of phages near the base of the carbuncles and furuncles in 6 patients led to a reduction in swelling, pain, and fever. |
| 1927 | India | V. cholerae | Cholera | This study was the first to use intravenous administration of bacteriophages by Asheshov in India. |
| 1929 | USA | Staphylococcus | Chronic furunculosis | Subcutaneous treatment of 208 patients showed 78% with no recurrent infections. |
| 1929 | USA | Staphylococcus | Septicemia | Remarkable success. |
| 1931 | Egypt | Yersinia pestis | Bubonic plague | In 1927, d’Herelle treated 4 cases of bubonic plague successfully by injecting bacteriophages into the buboes. |
| 1932 | USA | S. aureus | Meningitis | Remarkable success. |
| 1936 | USA | Staphylococcus | Staphylococcal bacteraemia | Relatively successful treatment in 100 of patients. |
| 1936 | France | Staphylococcus | Surgical infections | Cure of abscesses using polyvalent phages. |
| 1936 | Georgia | Salmonella typhi | Typhoid fever | Treatment of 21 patients resulted in a drop of 10.8% in mortality and 43.2% in complications. |
| 1936 | Georgia | Salmonella and Shigella | Acute colitis | All 43 patients with colitis were cured after treatment using “bacti-intesti-phage”. |
| 1936 | Georgia | S. typhi | Peritonitis caused by intestinal perforations in typhoid fever | Mortality was reduced from 85% to 20–35%. |
| 1942 | USA | Staphylococcus | Staphylococcal bacteraemia | Very positive results in the treatment of 500 patients. |
| 1946 | USA | Salmonella | Typhoid fever | The results were inconclusive; however, the positive results were astounding and encouraged them to continue the research. |
| 1949 | Canada | Salmonella | Typhoid fever | Several phage cocktails were used to treat 100 patients. The most successful one reduced the mortality rate from 20% to 2%. |
| 1968 | Russia | Salmonella and Shigella | Acute colitis | All 43 patients with colitis were cured after treatment using “bacti-intesti-phage”. |
| 1974 | Russia | Proteus, Staphylococcus, and Streptococcus | Peritonitis, osteomyelitis, lung abscesses, and postsurgical wound infections | Subcutaneous or through surgical wounds administration of phages in 236 patients resistant to antibiotics with a success rate of 92%. |
| 1976 | Russia | Abscessing pneumonia | A complex treatment was given to the patients, including intensive antibacterial therapy, immunotherapy, bacteriophage, protein preparations, vitamin therapy, and fresh blood transfusion. | |
| 1978 | Russia | E. coli and Proteus | Antibiotic-associated intestinal dysbiosis | A combination of phages and Bifidobacteria was used to restore the intestinal microbiota in 500 infants. |
| 1978 | Russia | E. coli, Proteus, Staphylococcus, and Streptococcus | Suppurative surgical infections | A comparison between commercial phage preparations and phages selected against bacterial strains isolated from patients was done. The selected phages were more effective in treating 60 patients. |
| 1979 | France | Enterobacter, Klebsiella, Proteus, Providencia, and S. aureus | Chronic orthopaedic infections | Seven cases of chronic orthopaedic infections were successfully treated with phages. |
| 1980 | Russia | E. coli, Proteus, Staphylococcus, and Streptococcus | Lung and pleural infections | Treatment of 45 patients using the combination of phages and antibiotics. |
| 1981 | Russia | E. coli and Proteus | Bacterial dysentery | A combination of phages and bifidobacteria was used to treat 59 immunosuppressed leukaemia patients. The treatment was reported to be more effective than antibiotics. |
| 1981–1986 | Poland | E. coli, Klebsiella, Pseudomonas, Salmonella, Shigella, and Staphylococcus | Gastrointestinal tract, skin, head, and neck infections | Phages were administered to over 1,000 patients in a series of studies. The success rates varied between 91% and 96%. |
| 1982 | Russia | Staphylococcus | Lung and pleural infections | Full recovery was reported in 82% of the patients treated with phages as opposed to 64% of patients treated with antibiotics. |
| 1984 | Russia | Shigella | Bacterial dysentery | The double-blinded study showed a 10-fold lower incidence of dysentery in those treated with phages. |
| 1984 | Russia | P. aeruginosa and S. aureus | - | A prophylactic mouthwash was administered to patients with acute leukemia. |
| 1986 | Poland | E. coli, Klebsiella, Proteus, Pseudomonas, and Staphylococcus | Bacterial monoinfections and polyinfections | The immunogenic effects of therapeutic phages were evaluated in 57 patients, showing an insignificant impact. |
| 1986 | Poland | Staphylococcus and several Gram-negative bacteria | Suppurative infections | During therapy, phages seemed to infiltrate the blood circulation and urinary tract. |
| 1987 | Poland | E. coli, Klebsiella, Proteus, Pseudomonas, and Staphylococcus | Suppurative skin infections | A success rate of 74% in 31 patients with chronically infected skin ulcers was observed upon phage administration. |
| 1988 | Russia | Pseudomonas and Staphylococcus | Post-surgical infections | Therapeutic phages were administered to 65 of 131 cancer patients, while the others received antibiotics. Phage therapy was successful in 82% of the patients in comparison to 61% success using antibiotics. |
| 1991 | Russia | Enterococcus, E. coli, P. aeruginosa, Proteus, Staphylococcus, and Streptococcus | Infectious allergosis | A total of 1,380 patients with infectious allergosis were treated using 3 different regimens: antibiotics, phages, and a mixture of both. The rates of success were 48%, 86%, and 83%, respectively. |
| 1992 | Russia | Klebsiella ozaenae, K. pneumoniae, and K. Rhinoscleromatis | Purulent inflammatory diseases | The administration of Klebsiella bacteriophages was successful in treating 109 patients with Klebsiella infections. |
| 1993 | Russia | Salmonella and Shigella | Bacterial dysentery and salmonellosis | A total of 1,646 children were successfully treated with phages and a combination of antibiotics and phages, where antibiotics alone were ineffective. |
| 1994 | Poland | E. coli | Recurrent subphrenic abscess | A case of recurrent subphrenic abscess caused by an antibiotic-resistant strain of E. coli was successfully treated with phages. |
| 1995 | Russia | E. coli, Proteus, and Staphylococcus | Inflammatory urogenital diseases | Adapted phages were used to treat 46 patients. The treatment was successful in 92% of the patients, while 84% showed bacterial clearance. |
| 1999 | Poland | K. pneumoniae | Cerebrospinal meningitis | Oral administration of a phage preparation successfully cleared bacteria from cerebrospinal fluid in a newborn. |
| 2001 | Russia | Proteus, Staphylococcus, and Streptococcus | Burn wounds | Pyophage treatment in patients with burn wounds reduced septic complications, had a 2-fold reduction of Staphylococci and Streptococci, a 1.5-fold reduction of Proteus, and a full reduction of E. coli. |
| 2002 | Republic of Georgia | E. coli, Proteus, Pseudomonas, and Staphylococcus | Ulcers and wounds | PhageBioDerm was administered and showed a 70% rate of success. |
| 2009 | UK | P. aeruginosa | Chronic otitis | Bacteriophage significantly reduced the numbers of P. aeruginosa in the phage-treated group in chronic otitis externa patients. |
| 2015 | France | S. aureus | Eye corneal abscess and interstitial keratitis | Bacteriophage eye drops with successful results after 6 months. |
| 2017 | Netherlands | S. aureus | Dermatoses | Successful treatment of chronic S. aureus with endolysin Staphefekt phage. |
| 2017 | Republic of Georgia | Staphylococci | Netherton syndrome | Successful treatment of manifestations of Netherton syndrome. |
| 2017 | Belgium | P. aeruginosa | P. aeruginosa septicaemia and acute kidney injury | Treatment of colistin-only-sensitive P. aeruginosa septicaemia. |
The Table is adapted with the permission of [36]. Copyright remains with the author(s) or their institution(s).
In 2017, phage therapy was performed on a patient with abdominal sepsis and diaphragmatic hernia in Belgium. The wound culture revealed the infection of multi-drug-resistant P. aeruginosa. Antibiotic therapy was stopped after observing acute kidney injury. The condition was very serious, with complications like a high C-reactive protein (CRP) level, low blood pressure, fever, and rapid heart rate. Then, bacteriophage therapy was opted for the treatment. Fifty microliters of purified bacteriophage cocktail BFC1 (two bacteriophages that showed activity against P. aeruginosa in vitro) was given to the patient intravenously, and the wound was also irrigated with the same. This treatment worked with negative blood culture, low CRP, and disappearance of fever. No side effects were noticed. The patient died after some months due to K. pneumoniae sepsis [131]. The authors also highlighted some cases of patients suffering from dermatoses caused by S. aureus and their successful treatment with the help of endolysin Staphefekt phage in the Netherlands [132]. Phage therapy against the antibiotic-resistant M. abscessus subsp. massiliense (strain GD01) infection in a human also suffering from cystic fibrosis was reported in 2019. 10,000 phages were considered that had been isolated from Mycobacterium smegmatis in the SEA-PHAGE program [128, 133]. Only 1,800 of them were genetically characterised. Further selection was done on the basis of infectivity against M. abscessus GD01. Only three (Muddy, JoeJ, and BPs) phages were selected for genetic modification using BRED. The repressor genes were removed either partially or totally in order to get the phage with only a lytic life cycle. After one month of bacteriophage mixture administration, the wound had healed in comparison to other wounds [128, 134].
The review mentioned key facts about normal commensals of the human body, drugs, and their characteristics, AMR, and various reasons that contribute to it. Bacteriophage, its characteristics, and how these characteristics are different from the drugs, with some advantages over the drugs. The key differences will help the reader to understand why bacteriophage is one of the suitable alternatives against bacterial infections. The author explained the bacteriophage characteristics and challenges in detail (bacteriophage replication, host specificity, and its mass production) and tried to suggest some of the important techniques that can be used for the respective desired modification. This review will help the reader to understand the key challenges in looking towards bacteriophages in place of antibiotics, and the reader will be able to make a conceptual picture of the modifications required. More investigation of this concept is important and revolutionary, not only for the healthcare industry. But for other sectors to decrease the effects of microbial growth, like food industries (by making bacteriophages specific for the bacterial communities that are mainly responsible for the spoilage and other gut-related diseases). In the food industry, this concept can replace chemical preservatives. In the agriculture sector, bacteriophages can be used against pests to protect the crops and to get a high yield.
AMR: antimicrobial resistance
BRED: bacteriophage recombineering of electroporated DNA
CRISPR: clustered regularly interspaced short palindromic repeat
CRP: C-reactive protein
HPLC: high-performance liquid chromatography
LPS: lipopolysaccharides
LVAD: left ventricular assist device
MIC: minimum inhibitory concentration
mTOR: mechanistic target of rapamycin
NMRC: Naval Medical Research Center
OmpC: outer membrane protein C
PCR: polymerase chain reaction
USA: United States of America
WHO: World Health Organisation
SP: Conceptualization, Writing—original draft, Writing—review & editing, Visualization. Medha S: Conceptualization, Writing—original draft. Manoj S: Writing—review & editing. All authors read and approved the submitted version.
The authors declare no conflicts of interest related to this work.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 182
Download: 7
Times Cited: 0