Review
Review

Affiliation:
Biotechnology Centre of Azores (CBA), University of the Azores, 9700-042 Azores, Portugal
Email: jorge.mr.medeiros@uac.pt
Explor Neurosci. 2025;4:100693 DOI: https://doi.org/10.37349/en.2025.100693
Received: January 25, 2025 Accepted: April 24, 2025 Published: June 06, 2025
Academic Editor: Ryszard Pluta, Medical University of Lublin, Poland
The article belongs to the special issue Alzheimer's Disease
Alzheimer’s disease, the main cause of dementia worldwide, is a slowly progressive neurodegenerative disorder. This disease involves a diversity of etiophatogenic processes as it is not only a genetic but also a biological and environmental disease. Owing to that complexity, nowadays there is no efficacious treatment for this disorder. The major Alzheimer’s disease clinical indications include extracellular senile plaques of amyloid-β protein, intracellular hyperphosphorylated τ neurofibrillary tangles, uncommon neuroinflammatory response, oxidative stress, and synaptic and neuronal dysfunction. The evaluation of the neuroprotective potential of new compounds is imperative. As natural products, like phenolic compounds, exhibit several bioactivities, it is urgent to test them and evaluate their inhibition of each clinical indication of Alzheimer’s disease. If phenolic compounds target more than one Alzheimer’s disease pathogenic mechanism (multi-target drug ligands), they will have the potential of becoming a leading Alzheimer’s disease treatment. Thus, this review analyzes, for each Alzheimer’s disease clinical indication, the scaffolds of several phenolic compounds leading to the highest activity with the objective to find phenolic compounds active against all the clinical indications. It was concluded that compounds presenting scaffolds like rugosin E or isocorilagin show potential in combating Alzheimer’s disease.
Alzheimer’s disease (AD) represents the most severe organic psychoneurological disease, comprising as much as 60% of dementia [1]. People affected by the disease usually present deficient cholinergic function, memory loss, loss of intellectual function, neuronal death, and behavioral disorders [2, 3]. Unfortunately, AD is not only a genetic disease but also a biological and environmental complex disease. That complexity leads to the lack of an effective treatment for AD [4, 5] and so the action against it has concerned mostly the reduction of the clinical indications (CIs) of the disease.
The major CIs of AD include extracellular plaques of Aβ-42 protein, intracellular neurofibrillary tangles (NFTs), uncommon neuroinflammatory response, oxidative stress, and synaptic and neuronal dysfunction [6–9]. The cleavage of the transmembrane amyloid precursor protein (APP) originates the formation of Aβ-42 plaques. By the amyloidogenic pathway APP is cleaved involving the action of two enzymes, β-secretase (BACE1) and γ-secretase. APP is cleaved by BACE1, resulting in two fragments, β-APP, and a longer peptide with 99 amino acids. The 99 amino acid fragments are now cleaved by γ-secretase into amyloidogenic peptides of varying length, including Aβ-42.
One way to combat AD is to prevent the appearance of the Aβ-42 plaques. Thus, one therapeutic strategy to combat AD is by the inhibition of the enzymes BACE1 and γ-secretase [8].
τ-Protein holds up the microtubules (MTs); however, when it is hyper-phosphorylated it aggregates itself and unties the MTs, which become destabilized. MTs are very important for the cytoskeleton in eukaryotic cells. MTs are always vibrating, alternating between growing and shrinking phases. A failure of these tuned actions of MTs originates the appearance of many neurodegenerative disorders, including AD. Therefore, the stabilization of MTs may potentially prevent AD progression. Another way to prevent the disease is to reduce hyper-phosphorylation of τ-protein, thereby avoiding MTs dysfunction. When τ-protein is hyper-phosphorylated it aggregates into paired helical and straight filaments that result in the formation of NFTs. As the phosphorylation of τ-protein results from an equilibrium between τ-kinase and phosphatase activities, kinase inhibitors restrain the processes of aggregation and the formation of NFTs. Thus, one of the key strategies to combat AD is the inhibition of the protein kinases used in the phosphorylation of τ-protein [10–23]. The main relevant protein kinases that interfere with τ-phosphorylation is glycogen synthetase kinase-3 beta (GSK3β) [24–26].
The development of AD may also be prevented by inhibiting the inflammatory response of microglial cells [27, 28]. The brain’s resident immune cells (microglia), under normal conditions, protect the brain from pathogens and help to maintain homeostasis of the tissues [29]. When insulted, the microglia cells adapt themselves, modifying their shapes, enabling their phagocytic functions to liberate a variety of such as nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin-1 (IL-1), IL-6, reactive oxygen species (ROS), prostaglandin E2 (PGE2), and cyclooxygenase-2 (COX-2). However, the accumulation of those proinflammatory factors results in damage and degeneration of the nearby neurons. Subsequently, the damaged neurons release certain immune substances, which increase the inflammatory neurotoxicity and cause irreversible neuroinflammation [30–35]. Thus, a potential therapeutic strategy for combating AD is also the use of agents for inhibiting the release of those proinflammatory factors.
Cognitive decline in AD patients is associated with the deficiency of the brain neurotransmitter acetylcholine (ACh). The enzymes acetylcholinesterase (AChE) and butyrylcholinesterase (BuChE) act by decomposing ACh and producing acetate and choline. When that happens, choline is taken up into the presynaptic neuron and carried out by the choline carriers, and so the signal transduction at the neuromuscular junction finishes rapidly [36]. Inhibition of AChE and BuChE prevents the breakdown of ACh, subsequently increasing its concentration and duration of action, which is clinically beneficial for AD patients. The use of AChE and BuChE inhibitors is widely used for the treatment of AD [37].
As described, AD has multiple pathogenic factors. So, the use of more than one pharmacological approach can be highly advantageous. One possible successful strategy might be the use of a single drug that can simultaneously hit multiple pathogenic factors, that is, a multitarget-directed ligand (MTDL).
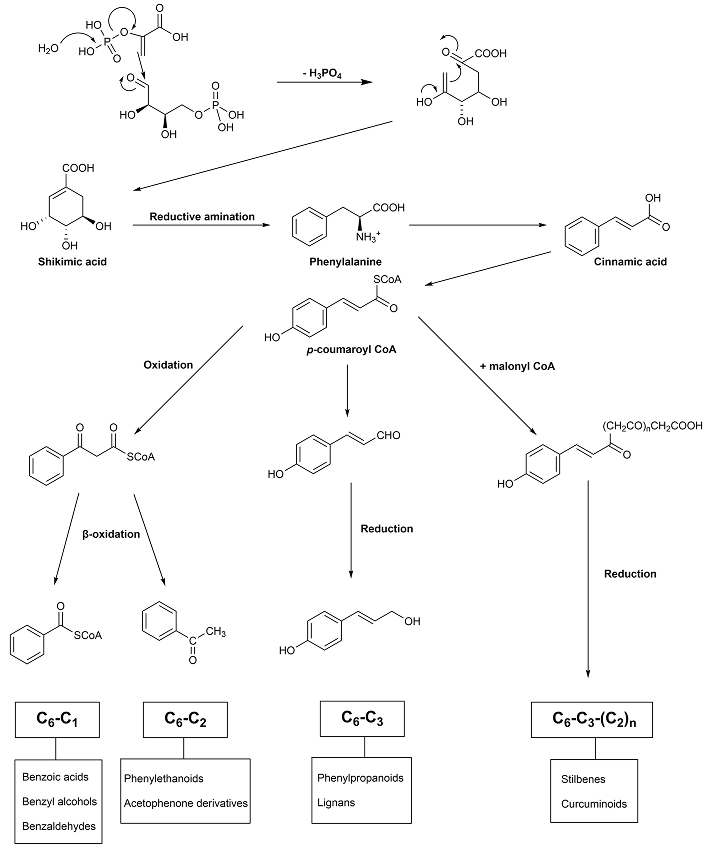
Among the several strategies that have been identified to combat AD, multi-target drug ligands have been shown to be an effective strategy for the treatment of this multifactorial disease, as compared to single-targeted agents and combined therapy [38]. Phenolic compounds are chemically defined as compounds containing one or more hydroxylated aromatic rings. They are derived from two main metabolic pathways—the shikimate and aceto-malonate pathways [39]. Except for flavonoids, all other plant phenolics are biosynthesized in plants through only the shikimate pathway [40–42]. As flavonoids were already reviewed [36], the phenolic compounds described in this review are biosynthesized by the shikimate pathway (Figure 1). The first step on this pathway consists of the formation of shikimic acid from phosphoenolpyruvic acid (PEP) and erythrose-4-phosphate through a series of steps.
This review focuses on the several phenolic compounds that have been analysed for their neuroprotective effects.
The load of amyloid plaques (Aβ-42) in the neuronal cells will be reduced by slowing the process.
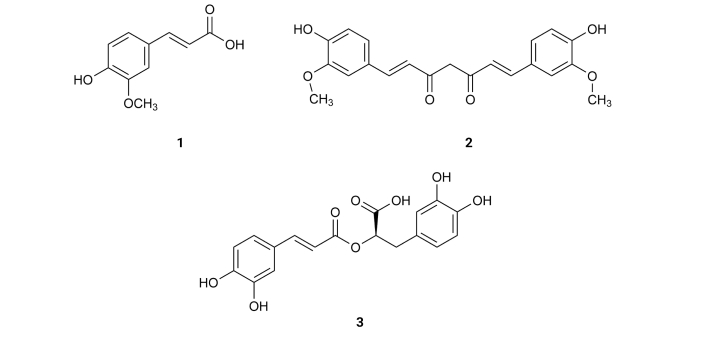
Ferulic acid (1), existing in Ferula communis [23], inhibits Aβ aggregation with an IC50 value of 9.30 µM [43] (Table 1). Curcumin (2), isolated from the rhizome of Curcuma longa, exhibits an IC50 value of 0.20 µM [44] (Table 1). Rosmarinic acid (3), derived from Lamiaceae, inhibits Aβ aggregation with an IC50 value of 20.3 µM [45] (Table 1). These results suggest that one of the structural requirements to express inhibitory activity on these compounds is the position of the phenolic hydroxyls, which should be in a para (p) position relative to the aliphatic side (Figure 2).
Phenolic compounds that inhibit clinical indications of Alzheimer’s disease
| No. | Compound | Mechanism | IC50(μM) | IC50(µmol/µmol AChE) | Eq. Trolox(mM) | Ref. |
|---|---|---|---|---|---|---|
| 1 | Ferulic acid | Aβ-42 | 9.3 | [43] | ||
| 2 | Curcumin | Aβ-42 | 0.2 | [44] | ||
| AChE | 58.08 | [108] | ||||
| 3 | Rosmarinic acid | Aβ-42 | 20.3 | [45] | ||
| τ-protein | 7.7 | [71] | ||||
| AChE | > 150 | [109] | ||||
| BuChE | 33.50 | [109] | ||||
| 4 | Resveratrol | Aβ-42 | 30.0 | [46] | ||
| AChE | 1.66 | [109] | ||||
| BuChE | 1.56 | [109] | ||||
| 5 | 3,4-Di-O-caffeoylquinic acid | Aβ-42 | 4.7 | [48] | ||
| BACE1 | 3.3 | [47] | ||||
| 6 | 3,5-Di-O-caffeoylquinic acid | Aβ-42 | 16.4 | [48] | ||
| 7 | 4,5-Di-O-caffeoylquinic acid | Aβ-42 | 0.1 | [48] | ||
| 8 | 1,4,5-Tri-O-caffeoylquinic acid | Aβ-42 | 2.2 | [48] | ||
| 9 | 3,4,5-Tri-O-caffeoylquinic acid | Aβ-42 | 0.3 | [48] | ||
| 10 | 3,4,5-Tri-O-caffeoylquinic acid methyl ester | Aβ-42 | 3.0 | [48] | ||
| 11 | Caffeic acid | Aβ-42 | 32.8 | [48] | ||
| NO | 40.0 | [75] | ||||
| AChE | 179.9 | [75] | ||||
| 12 | Chlorogenic acid | Aβ-42 | 92.9 | [48] | ||
| NO | 49.6 | [75] | ||||
| AChE | 410 | [102] | ||||
| 13 | Acteoside | Aβ-42 | 11.3 | [56] | ||
| AChE | 19.9 | [113] | ||||
| BuChE | 35.0 | [113] | ||||
| 14 | Cistanoside D | Aβ-42 | > 100 | [56] | ||
| 15 | Methylacteoside | Aβ-42 | > 100 | [56] | ||
| 16 | Oraposide | Aβ-42 | 8.2 | [56] | ||
| 17 | 3'''-O-methylcrenatoside | Aβ-42 | 28.4 | [56] | ||
| 18 | Methyloraposide | Aβ-42 | > 100 | [56] | ||
| 19 | Isoacteoside | Aβ-42 | 33.5 | [56] | ||
| 20 | Isocrenatoside | Aβ-42 | 27.4 | [56] | ||
| 21 | Hydroxytyrosol | Aβ-42 | 92.0 | [56] | ||
| 22 | Syringaldehyde | BACE1 | 34.90 | [58] | ||
| AChE | > 1,000 | [96] | ||||
| 23 | Benzoic acid | BACE1 | 309.30 | [58] | ||
| 24 | Phthalic acid | BACE1 | 133.40 | [58] | ||
| 25 | Urolithin B | BACE1 | 35.6 | [58] | ||
| 26 | (+)-Usnic acid | BACE1 | 43.1 | [58] | ||
| 27 | Esculetin | BACE1 | 7.67 | [59] | ||
| AChE | 6.13 | [59] | ||||
| BuChE | 8.66 | [59] | ||||
| 28 | Bergenin | BACE1 | > 400 | [60] | ||
| 29 | 11-O-p-hydroxybenzoylbergenin | BACE1 | 23.80 | [60] | ||
| 30 | 11-O-protocatechoylbergenin | BACE1 | 0.60 | [60] | ||
| 31 | 1,2,3-Trigalloyl glucopyranoside | BACE1 | 9.43 | [63] | ||
| 32 | Acetonyl geraniin | BACE1 | 0.71 | [63] | ||
| 33 | Helioscopinin A | BACE1 | 0.99 | [63] | ||
| 34 | Helioscopinin B | BACE1 | 0.41 | [63] | ||
| 35 | Furosin | BACE1 | 17.77 | [63] | ||
| 36 | Rugosin E | BACE1 | 0.06 | [63] | ||
| 37 | Euphorscopin | BACE1 | 2.50 | [63] | ||
| 38 | Jolkinin | BACE1 | 54.93 | [63] | ||
| 39 | 1-Galloyl glucopyranose | BACE1 | > 350 | [63] | ||
| 40 | 1,6-Digalloyl glucopyranose | BACE1 | > 350 | [63] | ||
| 41 | 2,6-Digalloyl glucopyranose | BACE1 | > 350 | [63] | ||
| 42 | 1,2,3,4,6-Pentagalloyl glucopyranose | BACE1 | > 350 | [63] | ||
| 43 | 1,2,6-Trigalloyl glucopyranose | BACE1 | > 350 | [63] | ||
| 44 | 2-Galloyl galactose | BACE1 | > 350 | [63] | ||
| 45 | Corilagin | BACE1 | > 350 | [63] | ||
| 46 | Elaeocarpusin | BACE1 | > 350 | [63] | ||
| 47 | Geraniin | BACE1 | > 350 | [63] | ||
| 48 | Helioscopin B | BACE1 | > 350 | [63] | ||
| 49 | 1,2,6-Trigalloyl allose | BACE1 | > 350 | [63] | ||
| 50 | 1,3,6-Trigalloyl allose | BACE1 | > 350 | [63] | ||
| 51 | 1,2,3,6-Tetragalloyl allose | BACE1 | > 350 | [63] | ||
| 52 | Bixanin | BACE1 | > 350 | [63] | ||
| 53 | 3-O-galloylshikimic acid | BACE1 | > 350 | [63] | ||
| 54 | 5-Pyrogallo-O-quinic acid | BACE1 | > 350 | [63] | ||
| 55 | α-Viniferin | BACE1 | > 350 | [63] | ||
| 56 | Schizandrin | BACE1 | > 350 | [63] | ||
| 57 | Gastrodin | GSK3β | NE | [73] | ||
| 58 | Salidroside | GSK3β | NE | [74] | ||
| 59 | Methylchlorogenic acid | NO | 34.9 | [75] | ||
| 60 | (E)-1,7-diphenylhept-4-en-3-one | SET-OH/H HAT-OOH | 149.8 | 0.28 | [76] | |
| AChE | 277.8 | [76] | ||||
| BuChE | NA | [76] | ||||
| 61 | 5-hydroxy-7-(4-hydroxy -3-methoxyphenyl)-1-phenylheptan-3-one | SET-OH/H HAT-OOH | 14.81 | 1.74 | [76] | |
| AChE | 190.7 | [76] | ||||
| BuChE | 252.0 | [76] | ||||
| 62 | (4Z,6E)-5-hydroxy-1,7-diphenylhepta-4,6-dien-3-one | SET-OH/H HAT-OOH | > 250 | 0.33 | [76] | |
| AChE | 194.5 | [76] | ||||
| BuChE | NA | [76] | ||||
| 63 | p-Hydroxycinnamic acid | SET-OH/H HAT-OOH | 29.94 | 3.20 | [76] | |
| AChE | 68.5 | [113] | ||||
| BuChE | > 100 | [113] | ||||
| 64 | p-Hydroxybenzoic acid | AChE | 150.6 | 6.36 | [77, 95] | |
| 65 | 2-Hydroxy-4-methoxybenzaldehyde | AChE | 47.0 | [78] | ||
| 66 | 4-Hydroxy-3-methoxybenzaldehyde | AChE | 37.0 | [78] | ||
| 67 | Methyl syringate | AChE | 5.50 | [95] | ||
| 68 | p-Hydroxyphenylpyruvic acid | AChE | 5.84 | [95] | ||
| 69 | Salicylic acid | AChE | 6.07 | [95] | ||
| 70 | p-Hydroxyphenylacetic acid | AChE | 6.24 | [95] | ||
| 71 | Homovanillic acid | AChE | 6.45 | [95] | ||
| 72 | Nordihydroguaiaretic acid | AChE | 6.47 | [95] | ||
| 73 | Protocatechuic acid | AChE | 6.50 | [95] | ||
| 74 | m-Hydroxybenzoic acid | AChE | 6.68 | [95] | ||
| 75 | Vanillic acid | AChE | 923 | 6.79 | [95, 102] | |
| 76 | Syringic acid | AChE | 6.96 | [95] | ||
| 77 | Homogentisic acid | AChE | 7.16 | [95] | ||
| 78 | Gentisic acid | AChE | 8.02 | [95] | ||
| 79 | Gallic acid | AChE | 9.32 | [95] | ||
| 80 | Ethyl-p-hydroxybenzoate | AChE | 31.38 | [95] | ||
| 81 | Ethyl vanillate | AChE | 34.19 | [95] | ||
| 82 | Dihydroconiferyl dihydro-p-coumarate | AChE | 357.9 | [96] | ||
| 83 | Gigantol | AChE | > 1,000 | [96] | ||
| 84 | Olivetol | AChE | 0.005 | [98] | ||
| BuChE | 0.006 | [98] | ||||
| 85 | 3,5-Dihydroxybenzoic acid | AChE | > 1,000 | [102] | ||
| 86 | Hydroquinone | AChE | 260 | [102] | ||
| 87 | Thymohydroquinone | AChE | 240.6 | [103, 104] | ||
| 88 | Carvacrol | AChE | 419.4 | [103, 104] | ||
| 89 | Thymol | AChE | > 1,000 | [103, 104] | ||
| 90 | 3,3'-Di-O-methylellagic acid | AChE | 141.6 | [105] | ||
| BuChE | 152.8 | [105] | ||||
| 91 | 3,3',4'-Tri-O-methylellagic acid-4-O-β-D-xylopyranoside | AChE | 128.6 | [105] | ||
| BuChE | > 200 | [105] | ||||
| 92 | 3,3',4'-Tri-O-methylellagic acid-4-O-β-D-glucopyranoside | AChE | 118.0 | [105] | ||
| BuChE | 148.5 | [105] | ||||
| 93 | 3,3'-Di-O-methylellagic acid-4-O-β-D-glucopyranoside | AChE | 129.1 | [105] | ||
| BuChE | 182.4 | [105] | ||||
| 94 | Anisacanthin | AChE | 0.09 | [106] | ||
| 95 | Pinoresinol | AChE | 0.292 | [106] | ||
| 96 | Epipinoresinol | AChE | 0.242 | [106] | ||
| 97 | Phillyrin | AChE | 0.279 | [106] | ||
| 98 | Pinoresinol-4-O-β-D-glucoside | AChE | 0.64 | [106] | ||
| 99 | Tannic acid | AChE | 0.12 | [107] | ||
| BuChE | 0.09 | [107] | ||||
| 100 | Salvianolic acid B | AChE | > 150 | [109] | ||
| BuChE | 14.6 | [109] | ||||
| 101 | Salvianolic acid A | AChE | > 150 | [109] | ||
| BuChE | 97.7 | [109] | ||||
| 102 | Carnosic acid | AChE | 95.8 | [109] | ||
| BuChE | 12.4 | [109] | ||||
| 103 | Carnosol | AChE | 33.7 | [109] | ||
| BuChE | 11.9 | [109] | ||||
| 104 | Danshensu salt | AChE | NA | [109] | ||
| BuChE | 109.8 | [109] | ||||
| 105 | Eugenol | AChE | 8.69 | [112] | ||
| BuChE | 8.86 | [112] | ||||
| 106 | Butylated hydroxyl toluene | AChE | 9.16 | [112] | ||
| BuChE | 9.00 | [112] | ||||
| 107 | Martinoside | AChE | > 100 | [113] | ||
| BuChE | > 100 | [113] | ||||
| 108 | Cinnamic acid | AChE | 78.5 | [113] | ||
| BuChE | > 100 | [113] | ||||
| 109 | Isoacteoside | AChE | 21.9 | [113] | ||
| BuChE | 29.7 | [113] | ||||
| 110 | Decaffeoylverbascoside | AChE | 16.1 | [113] | ||
| BuChE | 46.0 | [113] | ||||
| 111 | Lavandulifolioside | AChE | 301.0 | [114] | ||
| 112 | Lapathoside B | AChE | > 100 | [115] | ||
| BuChE | 10.9 | [115] | ||||
| 113 | Vanicoside B | AChE | 32.3 | [115] | ||
| BuChE | 7.5 | [115] | ||||
| 114 | Lapathoside A | AChE | 30.6 | [115] | ||
| BuChE | 2.7 | [115] | ||||
| 115 | Smilaside J | AChE | 56.0 | [115] | ||
| BuChE | 10.1 | [115] | ||||
| 116 | Smilaside G | AChE | > 100 | [115] | ||
| BuChE | 17.1 | [115] | ||||
| 117 | 4-O-caffeoyl quinic acid | AChE | 80.2 | [116] | ||
| 118 | 4,5-Di-O-caffeoyl quinic acid | AChE | 62.6 | [116] | ||
| 119 | Isocorilagin | AChE | 0.49 | [117] | ||
| BuChE | 4.20 | [117] |
NE: not evaluated; NA: not active
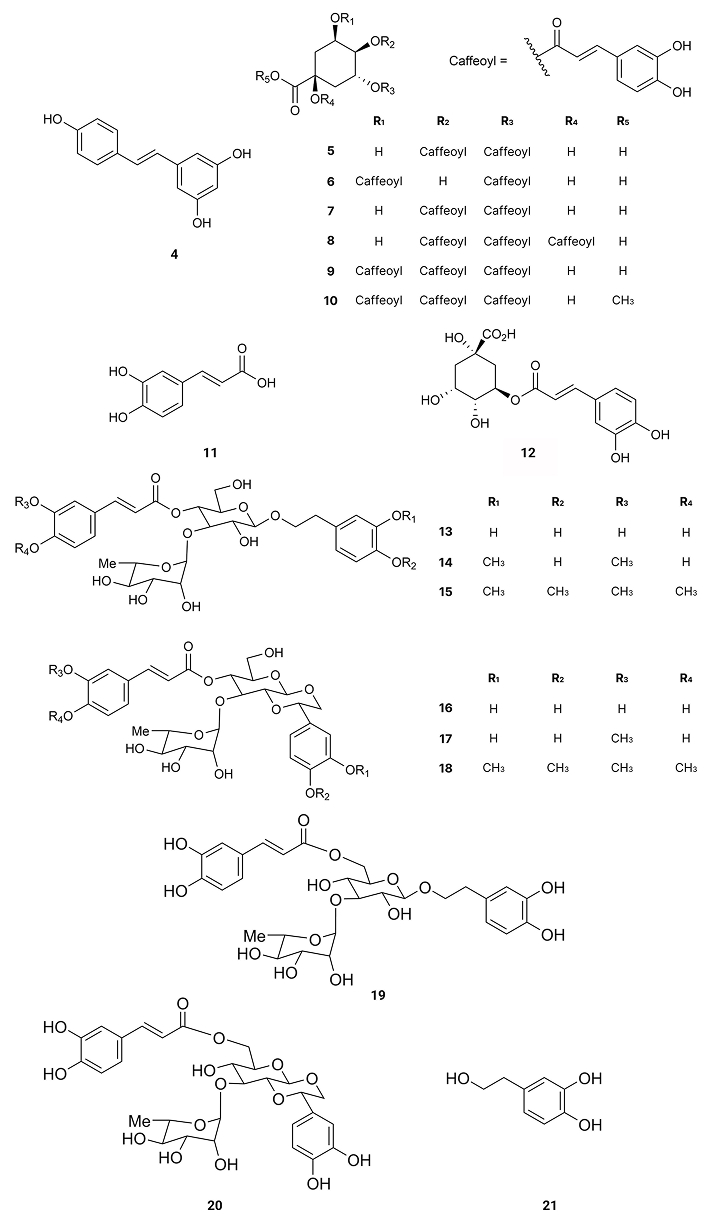
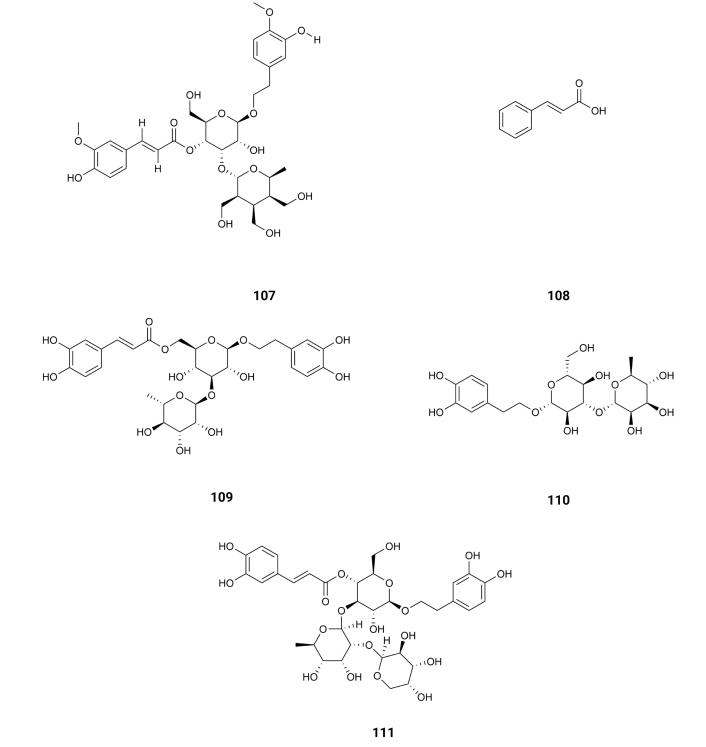
Resveratrol (4), existing in the skin of grapes, blueberries, raspberries, mulberries, and peanuts, promotes intracellular degradation of Aβ, via a mechanism that involves the proteasome, with an IC50 value of around 30.0 µM [46] (Table 1). 3,4-Di-O-caffeoylquinic acid (5) is present in Andrographis paniculata, which exists in peninsular India, Sri Lanka, as well as in different regions of Southeast Asia, China, America, the West Indies, and Christmas Island [47]. It presents inhibitory effects on Aβ-42 aggregation, exhibiting an IC50 value of 4.7 µM [48]. 3,5-Di-O-caffeoylquinic acid (6), isolated from Chrysanthemum spp. and Arctium spp. [49], 4,5-di-O-caffeoylquinic acid (7), isolated from Ipomoea batatas L. [50], 1,4,5-tri-O-caffeoylquinic acid (8), existing in Arnica montana and Arnica chamissonis [51], 3,4,5-tri-O-caffeoylquinic acid (9), isolated from Brasilian propolis [52] and 3,4,5-tri-O-caffeoylquinic acid methyl ester (10), existing in Lonicera japonica Thunb [53] also exhibit inhibitory activity on Aβ-42 aggregation with IC50 values of 16.4, 0.1, 2.2, 0.3 and 3.0 µM, respectively [48] (Table 1). On the other hand, compounds like caffeic acid (11) existing in Eucalyptus globulus [54] and chlorogenic acid (12) (Figure 3), found in the bamboo Phyllostachys edulis [55], were slightly active with IC50 values of 32.8 and 92.9 µM, respectively [48] (Table 1). From Orobanche minor were isolated acteoside (13), cistanoside D (14), methylacteoside (15), oraposide (16), 3'''-O-methylcrenatoside (17), methyloraposide (18), isoacteoside (19), isocrenatoside (20) and hydroxytyrosol (21) (Figure 3), which presented Aβ aggregation inhibitory activity with IC50 values of 11.3, > 100, > 100, 8.2, 28.4, > 100, 33.5, 27.4 and 92.0 µM, respectively [56] (Table 1).
These results show the importance of a caffeoyl group for the inhibitory activity on Aβ-42 aggregation. Indeed, by auto-oxidation, the catechol unit is susceptible to turn into an O-quinone, which might form a covalent bond with some residues of Aβ-42. Such modification may destabilize the β-sheet structure, turning it into amyloidogenic polypeptides. Thus, compounds with two or three catechol units like 4,5-di-O-caffeoylquinic acid (7), 3,4,5-tri-O-caffeoylquinic acid (9) or 3,4,5-tri-O-caffeoylquinic acid methyl ester (10) inhibit β-sheet formation of Aβ-42 while caffeic acid (11) and chlorogenic acid (12), with only one catechol unit, are slightly active [48] and methylacteoside (15) and methyloraposide (18) with no hydroxyl groups on the benzene rings are inactive. On the other hand, the presence of more than one caffeoyl group on the same side of the cyclohexane molecule decreases the inhibitory activity on Aβ-42 aggregation due to steric reasons.
Being BACE1, the enzyme involved in the rate-limiting step in the production of Aβ-42 plaques, the inhibition of this protease will reduce the load of the Aβ-42 plaques in the neuronal cells by slowing the process [57].
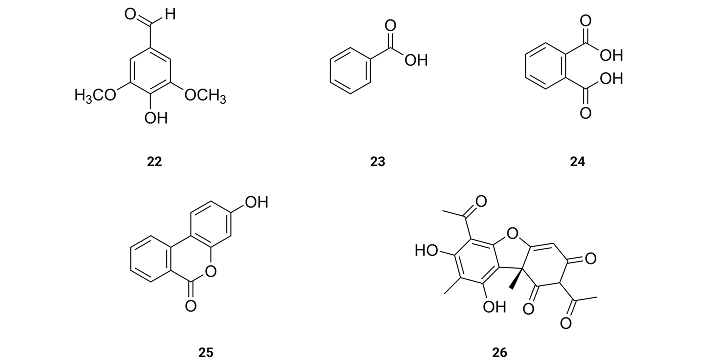
Syringaldehyde (22), benzoic acid (23), phthalic acid (24), urolithin B (25) and (+)-usnic acid (26) (Figure 4), isolated from the lichen Xanthoparmelia somloensisthe (Gyel.) Hale collected around Mongolian Shilajit, exhibit IC50 values for the inhibition of BACE1 of 34.9, 309.3, 133.4, 35.6 and 43.1 µM [58] (Table 1). Benzoic acid (23) and phthalic acid (24) with no hydroxyl group on the benzene rings are very weak inhibitors (IC50 > 100 µM). These results suggest that the presence of hydroxyl groups on the benzene ring is essential for the inhibition of BACE1.
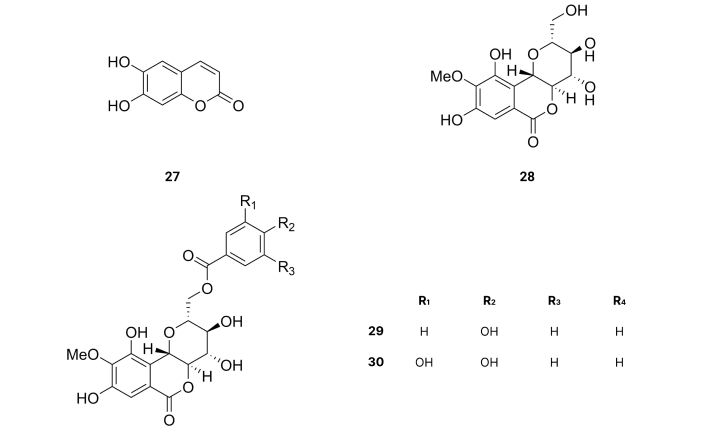
From the Chinese herbal medicine Fraxinus rhynchophylla Hance, esculetin (27) was extracted. The inhibition of the activities of BACE1 presents IC50 values of 7.67 µM [59] (Table 1). These results suggest that the presence of two hydroxyl groups at the ortho position of a benzene ring as well as being part of a very planar molecule increases the activity for the inhibition of BACE1.
3,4-Di-O-caffeoylquinic acid (5), isolated from Andrographis paniculata which exists in peninsular India, Sri Lanka, as well as, in different regions of Southeast Asia, China, America, the West Indies, and Christmas Island, exhibits an IC50 value of 3.30 µM [47] (Table 1) as BACE1 inhibitory activity.
Bergenin (28) isolated from the bark of Bergenia ligulate, Japan [60], 11-O-p-hydroxybenzoylbergenin (29), and 11-O-protocatechoylbergenin (30) (Figure 5), isolated from Bergenia ciliata (Haw) Sternb., Asia [61], present inhibitory activity of BACE1 with IC50 values of > 400, 23.8 and 0.6 µM [60] (Table 1), suggesting again that the presence of two hydroxyl groups at the ortho position of a benzene ring as 11-O-protocatechoylbergenin (30), as well as being part of a very planar group of a molecule like the benzoyl group, increases the activity for the inhibition of BACE1.
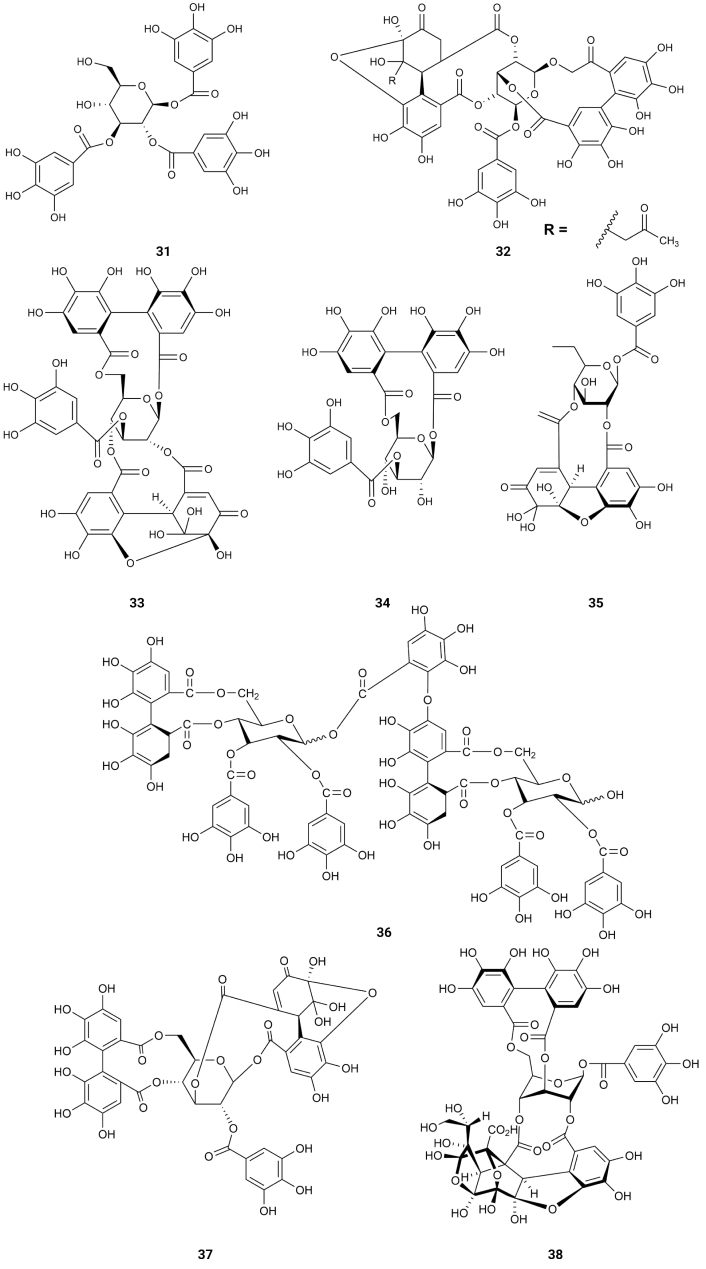
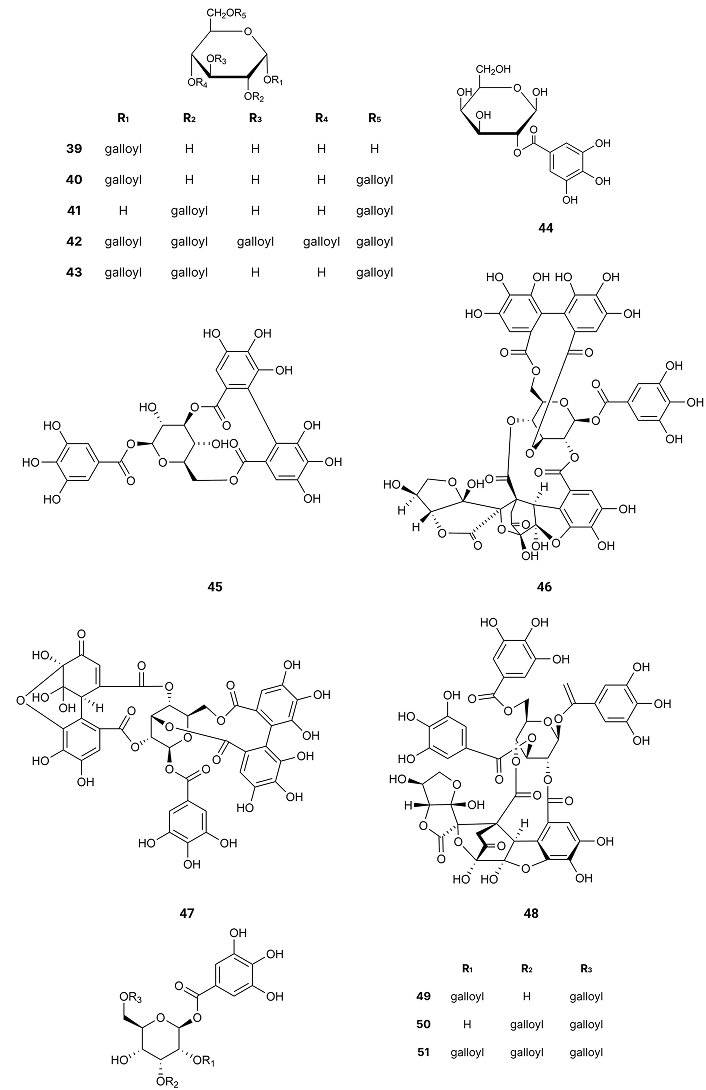
1,2,3-Trigalloyl glucopyranoside (31) isolated from Euphorbia prostrata, acetonyl geraniin (32) from Euphoria longana Lam., helioscopinin A (33) and helioscopinin B (34) from Euphorbia helioscopia L., furosin (35) from Erodium moschatum, rugosin E (36), from Euphorbia supina, euphorscopin (37) from Euphorbia helioscopia L. and jolkinin (38) (Figure 6) from Euphorbia jolkinii existing in Shangri-La, the Southwest China [62], exhibited strong inhibition of BACE1 with IC50 values of 9.43, 0.71, 0.99, 0.41, 17.77, 0.06, 2.50 and 54.93 µM [63] (Table 1). Rugosin E (36) was the most potent (IC50 = 0.06 µM) [63], which is the only compound with two 4,4',5,5',6,6'-hexahydroxydiphenoyl (HHDP) groups linked to the 4,6-positions of the glucopyranose core. 1-Galloyl glucopyranose (39), 1,6-digalloyl glucopyranose (40), 2,6-digalloyl glucopyranose (41) and 1,2,3,4,6-pentagalloyl glucopyranose (42), from Euphorbia helioscopia L. collected in Fukuoka, Japan [63]; 1,2,6-trigalloyl glucopyranose (43) from Euphorbia supina Rafin collected in Fukuoka, Japan [64]; 2-galloyl galactose (44), corilagin (45), elaeocarpusin (46), geraniin (47) and helioscopin B (48) from Euphorbia helioscopia L. collected in Fukuoka, Japan [62, 64]; 1,2,6-trigalloyl allose (49), 1,3,6-trigalloyl allose (50) and 1,2,3,6-tetragalloyl allose (51) from Euphorbia fischeriana, Japan [65] (Figure 7); Bixanin (52) existing in Macaranga sinensis (Baill.) Müll.Arg., China [66]; 3-O-galloylshikimic acid (53) existing in Arbutus unedo L. Mediterranean fringe [67]; 5-pyrogallo-O-quinic acid (54) existing in Camellia sinensis L., Taiwan, China [68]; α-viniferin (55) existing in Shorea ovalis Blume, Indonesia [69] and schizandrin (56) (Figure 8) existing in Schisandra chinensis Turcz. (Baill.), Korea, [70] are very weak inhibitors of BACE1 (IC50 > 350 µM) [63] (Table 1). These results suggest the importance of having HHDP groups linked to the 1,6- or 4,6-positions of the glucopyranose core. Having those linkages of the HHDP groups, which are very planar, there is no steric hindrance of the other groups of the molecule to the hydrogen bonds between the hydroxyl groups of the benzene rings and the active centres of BACE1. Comparing the activities of acetonyl geraniin (32) and geraniin (47) it can be concluded that the inhibitory activity of the former (IC50 = 0.71 µM) is due to the acetonyl group and not to the hydroxyl groups linked to benzene ring of the HHDP group.
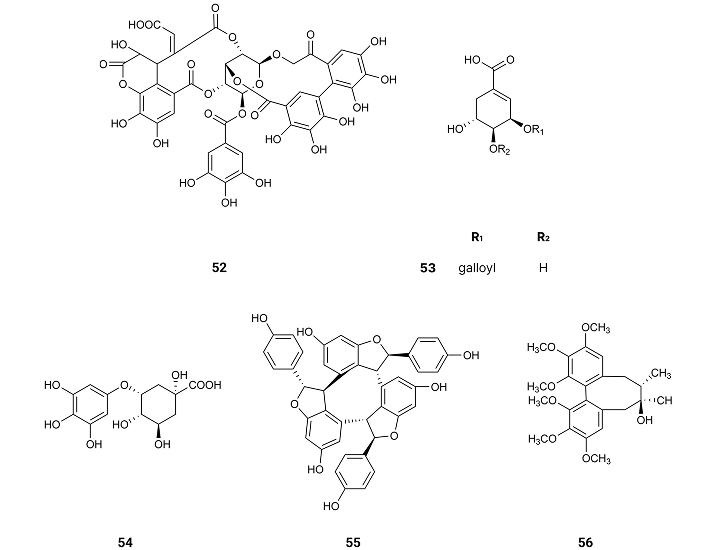
Rosmarinic acid (3) (Figure 2), isolated from Rosmarinus officinalis L., collected in Talca, Chileyet known., inhibits τ-protein in concentrations ranging from 10 µM to 100 µM, in a dose-dependent manner. This compound (3) exhibits an IC50 value of 7.7 µM [71] (Table 1). Rosmarinic acid (3) has two catechol moieties, suggesting that the presence of a catechol moiety might be important for the activity as an inhibitor of τ-protein. Indeed, τ-protein is a natural unstructured protein, and its exact structure is not yet known. However, the structure of the fibril-forming hexapeptide motif of τ-protein—306VQIVYK311—has been resolved by X-ray crystallography [71]. Molecular docking analysis concluded that rosmarinic acid (3) docking was done inside the cylindrical cavity, which is formed by paired 306VQIVYK311 β-sheets, in which compound (3) enters with no steric hindrance. As rosmarinic acid (3) has an elongated shape, aromatic and polar groups, and a negatively charged group, it might inhibit τ-aggregation by establishing chemical interactions with the fibril-forming hexapeptide VQIVYK. Thus, the aromatic rings of rosmarinic acid (3) stay packed against apolar side chains of Val309 and form several hydrogen bonds with glutamine and lysine side chain groups on both sides of the steric zipper. The most important interaction is the salt link between the carboxylate group of rosmarinic acid and two Lys311 ammonium ions from 306VQIVYK311 fibers [71].
As described above, another promising strategy to combat AD is the inhibition of protein kinases [10–23]. The most important protein kinase involved in τ-protein phosphorylation is GSK3β.
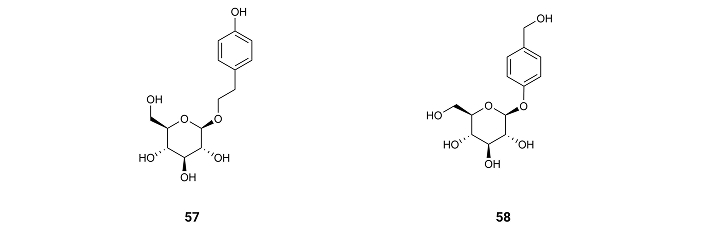
The compound gastrodin (57) (Figure 9), isolated from the orchid Gastrodia elata and from the rhizome of Galeola faberi [72], suppresses the activity of GSK3β in the brain of mice treated with a water solution of gastrodin (57) and administered it orally in a dosing volume of 10 mL/kg at doses of 150 mg/kg for nine consecutive months [73].
Salidroside (58) (Figure 9), a phenol glycoside compound found in plants of the Rhodiola genus, also decreases GSK3β activity after mice treatment by administering water solutions of the compound [74]. These results show the need for a phenol or benzyl group for the inhibition of GSK3β.
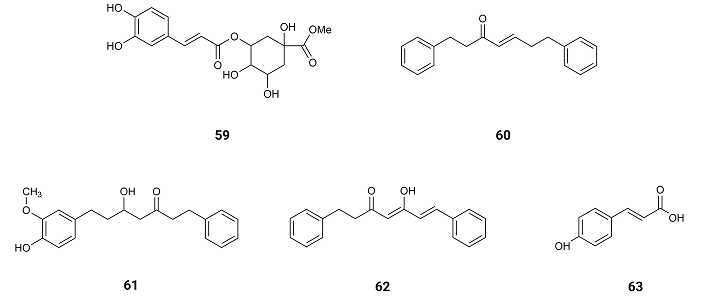
From the flowering aerial parts of Phagnalon saxatile (L.) Cass. which were collected, in Cefalù, Capo Playa (Sicily), Southern Italy, caffeic acid (11), chlorogenic acid (12), methylchlorogenic acid (59) (Figure 10), were isolated, exhibiting inhibitory effect against NO production with IC50 values of 40.0, 49.6 and 34.9 µM [75], respectively (Table 1).
(E)-1,7-diphenylhept-4-en-3-one (60), 5-hydroxy-7-(4-hydroxy-3-methoxyphenyl)-1-phenylheptan-3-one (61), (4Z,6E)-5-hydroxy-1,7-diphenylhepta-4,6-dien-3-one (62) and p-hydroxycinnamic acid (63) (Figure 10) isolated from Alpinia officinarum Hance rhizomes at Saudi Arabia, reduce the expression of ROS by showing activity on single electron transfer (SET) mechanisms suitable for both hydrophobic and hydrophilic compounds (SET-OH/H) or on hydrogen atom transfer (HAT) mechanisms as scavenging peroxyl radicles (HAT-OOR). The inhibition to produce free radicals by the SET-OH/H mechanism presents IC50 values of 149.8, 14.81, > 250 and 29.94 µM, respectively (Table 1). On the other hand, the antioxidant potential following the HAT-OOH mechanism of the compounds estimated as Trolox equivalents presented 0.28, 1.74, 0.33 and 3.20 mM TE/mM compound (Table 1). In both mechanisms 5-hydroxy-7-(4-hydroxy-3-methoxyphenyl)-1-phenylheptan-3-one (61) and p-hydroxycinnamic acid (63) showed the most powerful antioxidant effects [76], suggesting the need of an hydroxyl group on the para position for the inhibition of ROS production.
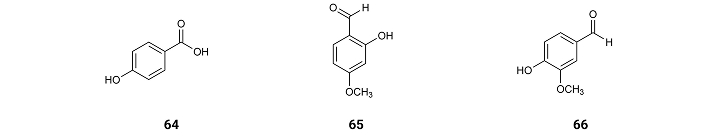
From Carthamus tinctorius L. flowers from Jeddah, Saudi Arabia, was isolated p-hydroxybenzoic acid (64), which displays inhibition of AChE with IC50 values of 150.6 µM [77]. 2-Hydroxy-4-methoxybenzaldehyde (MBALD) (65) from roots of Hemidesmus indicus H. indicus and 4-hydroxy-3-methoxybenzaldehyde (vanillin) (66) (Figure 11) from pod extracts of Vanilla planifolia exhibit inhibitory activity of AChE with IC50 values of 47.0 and 37.0 µM [78]. These results suggest that the aldehyde and the hydroxy group on MBALD (65) might be linked by intramolecular hydrogen bonding, not allowing any other hydrogen bonding with other molecules. When comparing the inhibition activities of p-hydroxybenzoic acid (64) and vanillin (66), it can be concluded that the methoxy group increases the inhibition activity by hydrophobic interactions with the enzyme.
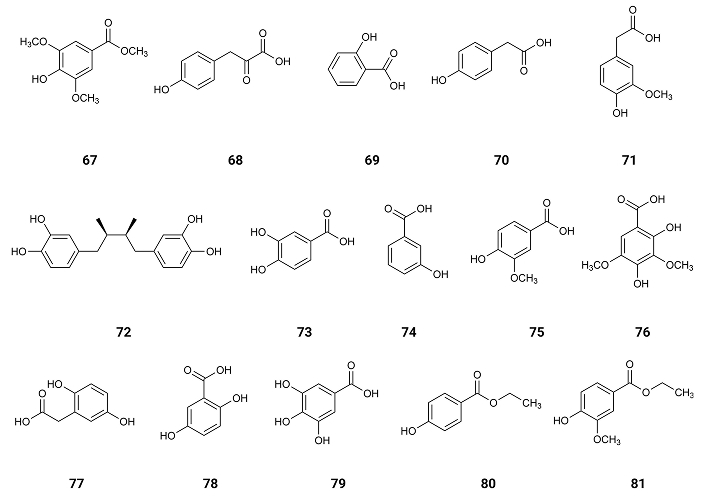
Methyl syringate (67) from Taraxacum formosanum, Taiwan, China [79]; p-hydroxyphenylpyruvic acid (68) from Agave angustifolia Haw, Mexico [80]; salicylic acid (69) from willow (Salix acmophylla Boiss.), found worldwide [81]; p-hydroxyphenylacetic acid (70) (Figure 12) found in olive oil [82]; p-hydroxybenzoic acid (64) found in Vitex agnus-castus, Mediterranean region [83]; homovanillic acid (71) found in Aloe greatheadii, Africa [84]; nordihydroguaiaretic acid (72), found in Larrea tridentata (Creosote bush), an abundant plant of Mexican and US-American deserts [85]; protocatechuic acid (73), found in Boswellia dalzielii, Africa [86]; m-hydroxybenzoic acid (74) which exists at Senna alata leaves, Brasil [87]; vanillic acid (75) found in Camellia sinensis (L.) Kuntze (green tea), China [88]; syringic acid (76) found in Ardisia elliptica Thunb, China [89]; homogentisic acid (77) isolated from the Indonesian fungus, Penicillium citrinum [90]; gentisic acid (78) (Figure 12) extracted from the wine of the berries of Vitis vinifera L. cv. Vidal, China [91]; gallic acid (79) existing in Quercus robur L., Poland [92]; ethyl p-hydroxybenzoate (80) existing in Tetragonia tetragonoides (Pall.) Kuntze, New Zealand [93]; and ethyl vanillate (81) existing in Sisymbrium officinale (L.) Scop., collected in the sub-Mediterranean region of South Croatia near Split [94] exhibit inhibition of AChE with IC50 values of 5.50, 5.84, 6.07, 6.24, 6.36, 6.45, 6.47, 6.50, 6.68, 6.79, 6.96, 7.16, 8.02, 9.32, 31.38 and 34.19 µmol/µmol AChE [95].
These results indicate that when O-hydroxybenzoic acid (salicylic acid) (69) is compared with the other isomers, p-hydroxybenzoic acid (64) and m-hydroxybenzoic acid (74), salicylic acid (69) shows the highest inhibition of AChE (IC50 = 6.07 µmol/µmol AChE) and p-hydroxybenzoic acid (64) shows a slightly lower inhibition (IC50 = 6.36 µmol/µmol AChE) [95].
It was noticed by Docking Simulation that both isomers were characterized by hydrogen bonds and hydrophobic interactions within the anionic binding site and the peripheral anionic site (PAS). m-Hydroxybenzoic acid (74) is less active than the other two isomers (IC50 = 6.68 µmol/µmol AChE). By Docking Simulation, it was realized that in this compound, the interactions with AChE were mainly through hydrogen bonds. These results suggest that the hydrophobic phenol-AChE interactions may be obstructed by the specific location of the hydroxyl groups [95].
Protocatechuic acid (3,4-dihydroxybenzoic acid) (73), with an additional hydroxyl group, presents an IC50 value of 6.50 µmol/µmol AChE. By Docking Simulation, it was realized that the interaction was almost identical to binding of the m-hydroxybenzoic acid (74) to AChE, with another hydroxyl group forming an additional hydrogen bond. However, as hydrogen bonding is a directed link, one of them is weaker than the other one. The isomer of 3,4-dihydroxybenzoic acid (73), 2,5-dihydroxybenzoic acid (gentisic acid) (78), exhibits a significantly lower activity (IC50 = 8.02 µmol/µmol AChE). Indeed, the interactions of gentisic acid (78) with AChE are very similar in nature to m-hydroxybenzoic acid (74). However, due to the presence of another hydroxyl group linked to carbon 2 of gentisic acid (78), the binding within the anionic binding site is decreased.
The presence of three hydroxyl groups in the aromatic ring of gallic acid (79) resulted in a weaker activity of the inhibitor (IC50 = 9.32 µmol/µmol AChE) compared to protocatechuic acid (73), with a similar pattern of interaction with the enzyme. In the case of the trihydroxy-derivative, there were interactions that weakened the binding of the complex (lower binding constant) related to the stress of the phenolic acid molecule.
The influence of the presence of methyl groups on the interactions with AChE was also analysed. Vanillic acid (4-hydroxy-3-methoxybenzoic) (75) showed weaker interactions of the hydrogen bond type than protocatechuic acid (73), due to the blocking of the hydroxyl group with a methyl substituent.
In the presence of two methyl groups, as in syringic acid (76) (Figure 12), hydrogen interactions prevailed in the complex when compared to hydrophobic interactions. However, in this case, the oxygen of the methoxy groups may also take part in a series of hydrophobic phenol-AChE interactions. As a result, the activity of syringic acid as an AChE inhibitor increases (IC50 = 6.96 µmol/µmol AChE) when compared to gallic acid (79) (Figure 12). However, the inhibitory activity of AChE of methyl derivative of syringic acid (76), methyl syringate (67), was very high (IC50 = 5.50 µmol/µmol AChE) suggesting that after the interaction of this compound with AChE the methyl group of the ester blocks the active centre of the enzyme completely and the hydrolysis of the ACh is impracticable [95].
When the interactions with AChE of ethyl derivatives like ethyl-p-hydroxybenzoate (80) and ethyl vanillate (81) (Figure 12) are analysed, they are characterized by a low binding constant. The interactions presented by p-hydroxybenzoic acid (64) are like the ones presented by the ethyl derivative ethyl-p-hydroxybenzoate (80). However, the higher stresses due to bond rotation, caused by the presence of the ethyl substituent and the loss of some interactions by hydrogen bonding, explain the difference in activity presented by these phenolic compounds. Vanillic acid (75) and its ethyl derivative ethyl vanillate (81) show similar interactions when analysed by calorimetry (ITC) and Docking Simulation, however, the binding constant was lower for ethyl vanillate (81) explaining the weaker inhibition of AChE (IC50 = 34.19 µmol/µmol AChE) presented by this compound suggesting the existence of some fewer and weaker hydrogen bonds between ethyl vanillate (81) and AChE. The derivatives of homohydroxyphenylacetic acids have also been analysed by calorimetry (ITC) and Docking Simulation. The activity of p-hydroxyphenylacetic acid (70) (IC50 = 6.24 µmol/µmol AChE) was like the one presented by p-hydroxybenzoic acid (64), but the type of interactions was different. The acetic derivative showed more hydrophobic interactions at the anionic binding site, and there were practically no repulsive forces. Similarly, homogentisic acid (77), when compared to gentisic acid (78), shows a higher activity as AChE inhibitor (IC50 = 7.16 µmol/µmol AChE), homovanillic acid (71) exhibits a higher activity than vanillic acid (75), p-hydroxyphenylpyruvic acid (68) presents a higher activity than p-hydroxyphenylacetic acid (70), which might be due to the greater number of significant interactions with AChE.
Nordihydroguaiaretic acid (72), a benzodiol dimer, due to its complex structure and the presence of two aromatic groups, may interact with AChE through many amino acids. However, due to the repulsive forces and stresses resulting from the rotation around the bonds, the interaction between the compound and AChE becomes weaker [95].
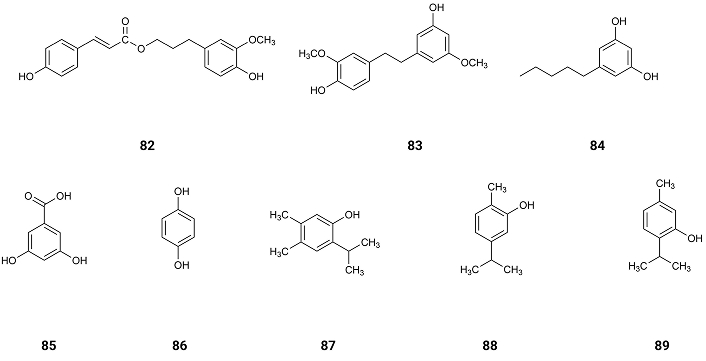
From Vanda roxburghii, found all over Bangladesh, syringaldehyde (22), dihydroconiferyl dihydro-p-coumarate (82) and gigantol (83) (Figure 13) were isolated. All of them were very weak or inactive as inhibitors of AChE with IC50 values of > 1,000, 357.9 and > 1,000 µM, respectively [96]. Syringaldehyde (22) is inactive. Indeed, as already mentioned, phenolic compounds with a structure like syringic acid (76) may have an alkyl group linked to the side chain, like methyl syringate (67), to block the active centre of the enzyme completely, and the hydrolysis of the AChE may be impossible [95].
From the Chinese herbal medicine Fraxinus rhynchophylla Hance, esculetin (27) was extracted. The inhibition of the activities of AChE and BuChE presents IC50 values of 6.13 and 8.66, respectively [59]. These results suggest that the presence of the two hydroxyl groups at the ortho position, as well as being an aglycone with no sugar moieties, makes the molecule very active.
Olivetol (84) (Figure 13), found in several Parmeliaceae lichens like Hypogymnia physodes, Evernia prunastri and Parmelia sulcata, in Serbia [97], presents activity as an inhibitor of AChE and BuChE with IC50 values of 0.005 and 0.006 µM, respectively [98]. As already mentioned above when two hydroxyl groups are on meta (m) substitution the interactions with AChE are mainly through hydrogen bonds, which on this compound are very strong as the two hydroxyl groups are well separated and the side chain, being aliphatic, where carbons are linked only by a sigma bond (σ), allows the entrance of the compound in the deep gorge of AChE.
Comparing phenolic compounds like 3,5-dihydroxybenzoic acid (85), present in Artemisia vulgaris L. and Artemisia alba Turra [99], vanillic acid (75), found in the roots of Angelica sinensis [100], hydroquinone (86) (Figure 13), found in Agaricus hondensis mushrooms [101] and chlorogenic acid (12), found in the bamboo Phyllostachys edulis [55], present weak inhibitory activity of AChE with IC50 values of > 1,000, 923, 260 and 410 µM, respectively [102], suggesting that the inhibitory activity of AChE depends on the number of hydroxyl groups and their distribution on the ring as well as of the aliphatic side chain.
In Thymus vulgaris L. of central Dalmatia, Croatia, several phenolic compounds as thymohydroquinone (87), carvacrol (88), and thymol (89) (Figure 13) which show some or no inhibitory activity of AChE. The IC50 values of them are 240.6, 419.4 and > 1,000 µM [103, 104] (Table 1). These results show again that the number of hydroxyl groups is important for the inhibition activity of AChE and their para position on the ring, as well as the steric effect of the isopropyl group.
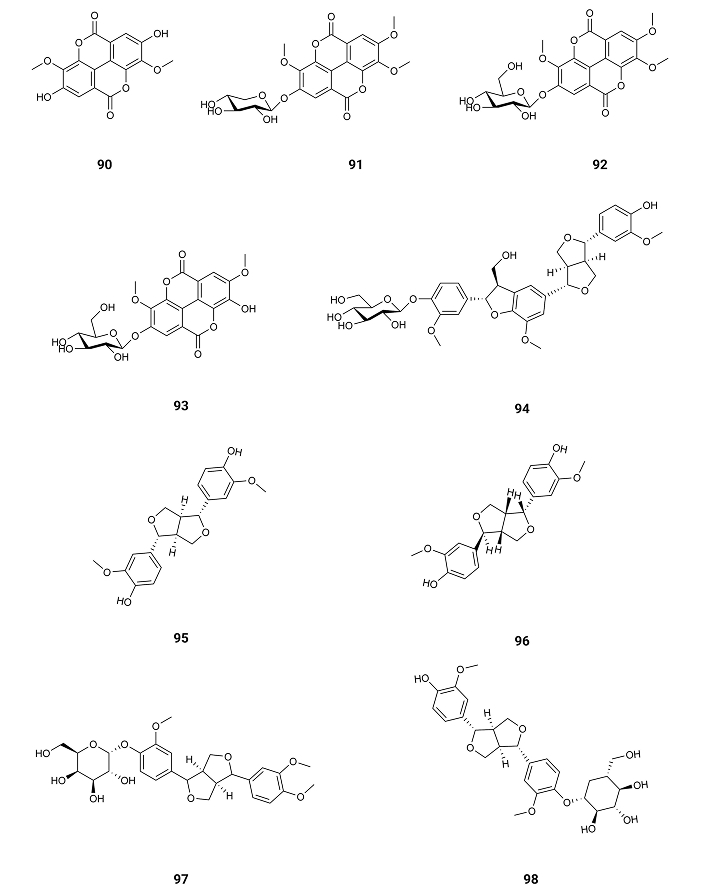
3,3'-Di-O-methylellagic acid (90), 3,3',4'-tri-O-methylellagic acid-4-O-β-D-xylopyranoside (91), 3,3',4'-tri-O-methylellagic acid-4-O-β-D-glucopyranoside (92), and 3,3'-di-O-methylellagic acid-4-O-β-D-glucopyranoside (93) (Figure 14) exist in Terminalia macrocarpa (Combretaceae) plant collected in Ngaoundere, Cameroon. The inhibitory activity of AChE exhibits IC50 values of 141.6, 128.6, 118.0, and 129.1 µM. Against BuChE, the IC50 values are 152.8, > 200, 148.5, and 182.4 µM [105] (Table 1). Therefore, the presence of a sugar moiety doesn’t change the inhibition of AChE for these types of compounds.
From fresh aerial parts of Anisacanthus virgularis (Salisb.) Nees gathered from Giza, Egypt, were extracted five furofuranoid-type lignans: anisacanthin (94), pinoresinol (95), epipinoresinol (96), phillyrin (97) and pinoresinol-4-O-β-D-glucoside (98) (Figure 14). These compounds show AChE inhibition with IC50 values of 0.09, 0.29, 0.24, 0.28, and 0.64 µM [106] (Table 1). All these molecules are active as inhibitors of AChE suggesting that the activity is due again to the aliphatic side chain, exhibiting anisacanthin (94) a higher activity as it is longer and more flexible the allowing the entrance of the compound in the wide and deep gorge of AChE.
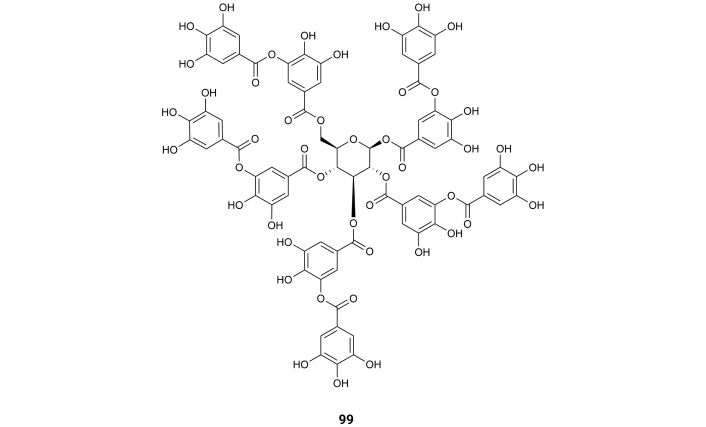
Tannic acid (penta‐m‐digalloyl‐glucose) (99) (Figure 15) is a natural polyphenolic compound found in some galls of Quercus species, Rhus species, beverages (red wine, tea, coffee), fruits, and vegetables. It presents inhibitory activity of AChE and BuChE with IC50 values of 0.12 and 0.09, respectively [107] (Table 1), suggesting that the several gallic acid moieties might interact with the enzyme AChE.
Curcumin (2) isolated from Curcuma longa, Ásia, presents inhibition of AChE with an IC50 value of 58.08 µM [108] (Table 1). Suggesting that, as the molecule is very planar, it has difficulty in entering the gorge of the enzyme.
(E)-1,7-diphenylhept-4-en-3-one (60), 5-hydroxy-7-(4-hydroxy-3-methoxyphenyl)-1-phenylheptan-3-one (61) and (4Z,6E)-5-hydroxy-1,7-diphenylhepta-4,6-dien-3-one (62) isolated from the rhizomes Alpinia officinarum Hance rhizomes, Saudi Arabia, exhibit inhibition of AChE with IC50 values of 277.8, 190.7, and 194.5 µM, respectively [76]. 5-hydroxy-7-(4-hydroxy-3-methoxyphenyl)-1-phenylheptan-3-one (61) also presented activity against BuChE with IC50 values of 252.0 µM. All three other compounds were inactive against BuChE [76]. These results suggest that the presence of one hydroxyl group in a benzene ring and a flexible side chain increases the binding capacity of phenolic compounds to AChE and BuChE.
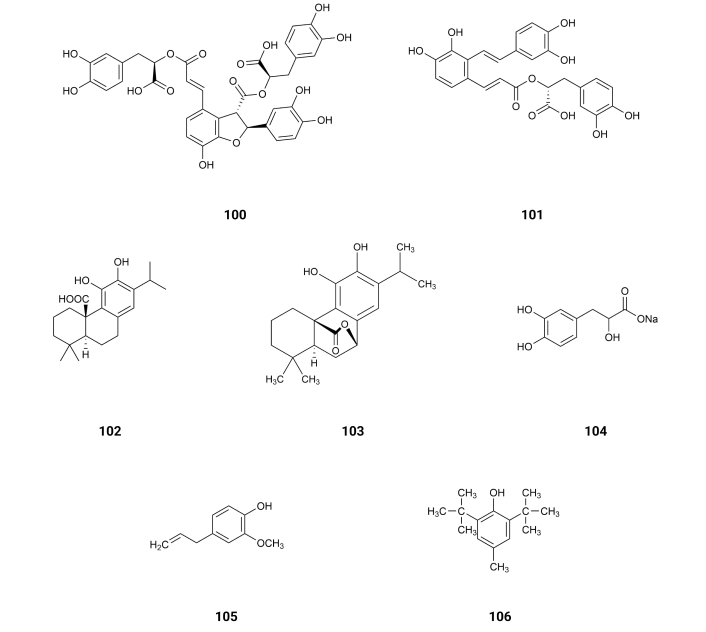
From Salvia L. species (Lamiaceae) existing at Asia [109], rosmarinic acid (3), salvianolic acid B (100), salvianolic acid A (101), carnosic acid (102), carnosol (103) and danshensu salt (104) (Figure 16) were extracted. The inhibition of AChE exhibits IC50 values of > 150, > 150, > 150, 95.8, 33.7, and >>150 (not active) µM [109] (Table 1), respectively. These compounds for the inhibition of BuChE exhibit IC50 values of 33.5, 14.6, 97.7, 12.4, 11.9, and 109.8 µM [109] (Table 1), respectively. It is interesting to notice that only carnosic acid (102) and carnosol (103) present two hydroxyl groups in an ortho-position. As already mentioned, by Docking Simulation was realized that the interaction between AChE and phenolic compounds with ortho hydroxyl groups is characterized by strong hydrogen bonds and hydrophobic interactions within the anionic binding site and PAS, explaining the increase of activity of these compounds for the inhibition of AChE.
Eugenol (105), existing in Cinnamomum zeylanicum Blume, India [110], and butylated hydroxyl toluene (106) (Figure 16), existing in the green algae Botryococcus braunii, Taiwan, China [111], exhibit activity as inhibitors of AChE with IC50 values of 8.69 and 9.16 µM, respectively. They also exhibit inhibition of BuChE with IC50 values of 8.86 and 9.00 µM, respectively [112]. These results suggest that the bulky t-butyl groups at the ortho-position relative to the hydroxyl group don’t prevent the hydrogen bonding of the hydroxyl group on butylated hydroxyl toluene (106).
From the dried roots of Harpagophytum procumbens (Pedaliaceae), Africa, were isolated martinoside (107), cinnamic acid (108), acteoside (13), isoacteoside (109), decaffeoylverbascoside (110) (Figure 17), and p-hydroxycinnamic acid (63) (Figure 10). They exhibit AChE inhibitory activity with IC50 values of > 100, 78.5, 19.9, 21.9, 16.1, and 68.5 µM [113], respectively. The inhibition of BuChE presents IC50 values of > 100, > 100, 35.0, 29.7, 46.0, and > 100 µM [113] (Table 1). It is interesting to compare the inhibition of AChE of martinoside (107) (IC50 > 100 µM) with the ones presented by acteoside (13) (IC50 = 19.9 µM), isoacteoside (109) (IC50 = 21.9 µM) and decaffeoylverbascoside (110) (IC50 = 16.1 µM). As already mentioned, the interaction is almost identical to the binding of the hydroxyl group at m-position to AChE with another hydroxyl group at p-position, forming an additional hydrogen bond with Glu199.
Lavandulifolioside (111) (Figure 17) isolated from Leonurus japonicus L. (Lamiaceae), Korea, presents inhibitory activity of AChE with an IC50 value of 301 µM [114]. The hydrolysate of lavandulifolioside (111), caffeic acid (11) (Figure 3), exhibits AChE inhibitory activity with IC50 value of 179.9 µM [75] suggesting that lavandulifolioside (111) with four hydroxyl groups linked to benzene rings cannot use all of them to link to the enzyme by steric reasons.
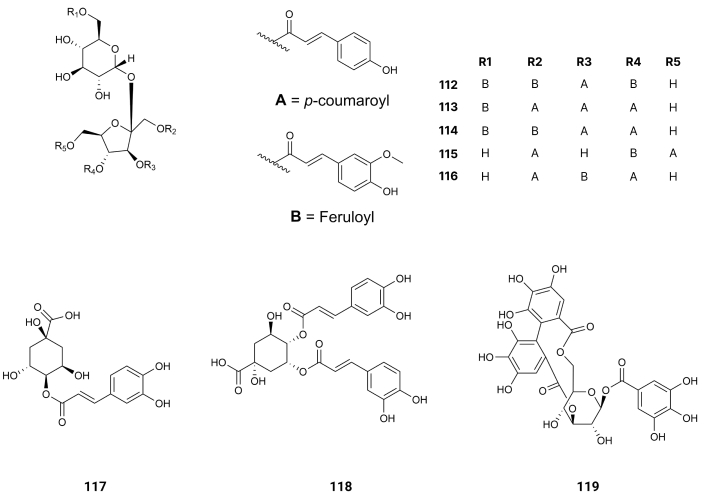
From Fallopia dentatoalata (Fr. Schm.) Holub, China, Lapathoside B (112), Vanicoside B (113), Lapathoside A (114), Smilaside J (115), and Smilaside G (116) (Figure 18) were extracted. They display inhibition of AChE with IC50 values of > 100, 32.3, 30.6, 56,0, and > 100 µM [115] (Table 1). The inhibition of BuChE exhibits IC50 values of 10.9, 7.5, 2.7, 10.1, and 17.1 µM [115] (Table 1). These results suggest that the feruloyl group produces on the side groups steric effects hampering the formation of hydrogen bonds on the side substituents.
4-O-Caffeoyl quinic acid (117) and 4,5-di-O-caffeoyl quinic acid (118) (Figure 18) were isolated from Acanthopanax henryi Harms, China. They display inhibition of AChE with IC50 values of 80.2 and 62.6 µM [116] (Table 1). The latter, having two more hydroxyl groups linked to benzene rings, is more active; however, the increase in activity is not proportional to the increase in hydroxyl groups of the benzene rings.
From Phyllanthus niruri leaves, Malaysia, isocorilagin (119) (Figure 18), this compound presents inhibition of AChE and BuChE with IC50 values of 0.49 and 4.20 μM, respectively [117]. Molecular docking analysis revealed that this compound, by the formation of hydrogen bonds with residues at the entrance of the AChE active site, effectively blocks it. With BuChE, the compound completely docked inside and occupied the active site of the enzyme [117].
Some phenolic compounds have been shown to be potential anti-Alzheimer agents. The most active phenolic compound against the BACE1 already tested is rugosin E (36) (IC50 = 0.06 µM) [63]. It is the only compound tested, until now, as an inhibitor of BACE1 with two 4,4',5,5',6,6'-HHDP groups linked to the 4,6-positions of a glucopyranose core. This result suggests the importance of having HHDP groups linked to the 1,6- or 4,6-positions of taglucopyranose core. Having those linkages of the HHDP groups, which are very planar, there is no steric hindrance of the other groups of the molecule to the hydrogen bonds between the hydroxyl groups of the benzene rings and the active centres of BACE1.
The compounds gastrodin (57) and salidroside (58) are the only compounds that presented activity against GSK3β. No other ones were studied yet however, it should be noted the presence of a phenoxy group on the former compound (57), which is at the para-position relative to the side chain of this compound C6–C1. The latter (58) also presents a hydroxy group at para-position relative to the side chain of this compound C6–C2, suggesting the need to have an oxygen atom linked to the benzene ring at para-position so that a phenolic compound can be active against GSK3β.
Concerning the inhibition of the pro-inflammatory factors 5-hydroxy-7-(4-hydroxy-3-methoxyphenyl)-1-phenylheptan-3-one (61) and p-hydroxycinnamic acid (63) showed the most powerful antioxidant effects [76] on both mechanisms—SET-OH/H and HAT-OOR—suggesting the need of an hydroxyl group on the para-position of phenolic compounds C6–C3 for the inhibition of ROS production.
For the inhibition of AChE, olivetol (84) presents the most powerful inhibition of AChE, suggesting the need for two hydroxyl groups on meta (m) substitution and an aliphatic side chain allowing the entrance of the compound in the deep gorge of AChE. On the other hand, comparing the inhibition of AChE of syringic acid (76) and methyl syringate (67) there is a surprising high increase of activity when the last compound is examined, suggesting that the methyl group of the ester blocks the active center of the enzyme completely and the hydrolysis of the ACh is impracticable. When comparing 4-O-caffeoyl quinic acid (117) and 4,5-di-O-caffeoyl quinic acid (118), the latter, having two units of the former, should have an enormous increase in activity as an inhibitor of AChE. Indeed, that increase is diminute. Thus, it is more convenient to have one phenolic compound than a molecule with several phenolic units. However, it is not the case for molecules like isocorilagin (119). Indeed, for this polyphenols compound, the formation of hydrogen bonds with residues at the entrance of the AChE active site blocks it completely.
The results discussed until now only concern the interaction of one phenolic compound with the active site of one enzyme. However, as AD is such a complex disease involving several mechanisms that may work together through interaction between genetic, molecular, and cellular events, one possible successful strategy might be treating AD with a multidrug combination in a more causally directed manner—multitarget therapy [118, 119]. This therapy can be achieved in two ways. One of them, called combination therapy, uses a drug cocktail, where each drug has an active component for the inhibition of one of the mechanisms of AD. This approach to therapy is associated with high risk drug-drug interactions. The other approach is called MTDL, where only one active ingredient is administered [120]. Using the MTDL approach, the risk of interaction between the several drugs of the combination therapy is eliminated, and the pharmacokinetic and pharmacodynamic properties are simplified with one single agent. Unfortunately, the data for every compound analysed is incomplete. There is no compound for which data exists concerning its activity against every CI. Analyzing the scaffolds of several phenolic compounds, which inhibit one of the mechanisms of AD, it is concluded that all of those mechanisms should be inhibited mostly by compounds presenting a scaffold like rugosin E (36) or isocorilagin (119).
ACh: acetylcholine
AChE: acetylcholinesterase
AD: Alzheimer’s disease
APP: amyloid precursor protein
BuChE: butyrylcholinesterase
CIs: clinical indications
GSK3β: glycogen synthetase kinase-3 beta
HAT: hydrogen atom transfer
HHDP: hexahydroxydiphenoyl
IL-1: interleukin-1
MTDL: multitarget-directed ligand
MTs: microtubules
NFTs: neurofibrillary tangles
PAS: peripheral anionic site
ROS: reactive oxygen species
SET: single electron transfer
JM: Writing—original draft, Writing—review & editing, Visualization, Conceptualization.
The author declares no conflict of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
© The Author(s) 2025.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2025. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

Janusz Wiesław Błaszczyk
Tatsushi Yuri ... Hisashi Nojima
Priyanka Sengupta ... Debashis Mukhopadhyay
Danqing Xiao, Chen Zhang
Carlos Gutierrez-Merino
Julius Mulumba ... Yong Yang
Felipe P. Perez ... Maher Rizkalla
Ezra C. Holston
Ryszard Pluta