 Original Article
Original Article

Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
2Saint Petersburg Research Institute of Phthisiopulmonology, Saint Petersburg 194064, Russia
ORCID: https://orcid.org/0000-0001-6359-0026
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
3Chair of Internal Medicine, North-Western State Medical University n. a. I. I. Mechnikov, Saint Petersburg 191015, Russia
4L.G. Sokolov North-Western District Scientific and Clinical Centre, Saint Petersburg 194291, Russia
ORCID: https://orcid.org/0000-0002-6815-444X
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
4L.G. Sokolov North-Western District Scientific and Clinical Centre, Saint Petersburg 194291, Russia
ORCID: https://orcid.org/0000-0002-7954-8567
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
5Saint Petersburg Research Institute of Phthisiopulmonology, Saint Petersburg 191036, Russia
ORCID: https://orcid.org/0000-0003-1174-7698
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
ORCID: https://orcid.org/0000-0003-1843-2808
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
ORCID: https://orcid.org/0000-0002-0807-1538
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
6Department of Pathophysiology with the Course of Immunopathology, Saint Petersburg State Pediatric Medical University, Saint Petersburg 194100, Russia
ORCID: https://orcid.org/0000-0001-9690-0043
Affiliation:
7I.M. Sechenov Institute of Evolutionary Physiology and Biochemistry of the Russian Academy of Sciences (IEPHB RAS), Saint Petersburg 194223, Russia
ORCID: https://orcid.org/0000-0002-2723-4590
Affiliation:
1Department of Pathology, Laboratory of the Microangiopathic Mechanisms of Atherogenesis, Saint Petersburg State University, Saint Petersburg 199034, Russia
Email: muslimbek.normatov@gmail.com
ORCID: https://orcid.org/0000-0002-1307-0513
Explor Immunol. 2026;6:1003257 DOI: https://doi.org/10.37349/ei.2026.1003257
Received: July 15, 2025 Accepted: April 24, 2026 Published: July 01, 2026
Academic Editor: Diego A. Bonilla, Dynamical Business & Science Society-DBSS International SAS, Colombia
Aim: This study explores the pathogenesis of atherosclerosis, as well as the comorbidity of atherosclerosis and coronavirus infection. The objective of the article is to provide a rationale for the potential involvement of an autoimmune component in the observed comorbidity between these two conditions.
Methods: The research utilized bioinformatic and laboratory techniques. The bioinformatic approach involved selecting 30 human autoantigens, either proven or hypothesized to participate in atherogenesis and/or arteritides, as well as the most immunogenic proteins from human coronavirus antigens. To identify shared minimal immunogenic determinants (pentapeptides) between human autoantigens and coronavirus antigens, the proprietary “Alignmentaj” program was employed. Antibody levels against apolipoprotein B-100 (ApoB-100) and Proteinase 3 (PR3) were measured using enzyme-linked immunosorbent assay (ELISA) in patients with cardiovascular disease following coronavirus disease 2019 (COVID-19). Post-mortem tissue samples were subjected to standard morphological examination, supplemented with immunohistochemical methods for pathomorphological analysis.
Results: The analysis revealed that the spike protein of human coronaviruses exhibits the highest concentration of pentapeptides similar to those found in atherogenesis-associated proteins. Notably, ApoB-100 shared the greatest number of peptides. Post-COVID cardiovascular patients showed varied anti-ApoB antibody and anti-PR3 antibody levels and potential complications. Pathomorphological examination of the aorta and coronary arteries of patients who died from atherothrombotic complications following COVID-19 infection revealed signs of autoimmune inflammation. Lymphocytic infiltration, consisting of T-cells and B-cells, as well as indications of vasa vasorum vasculitis, were observed in areas of unstable aortic atherosclerotic plaques and in the adventitia of coronary arteries.
Conclusions: The identified sequence homology between human coronavirus antigens and human autoantigens, together with laboratory data obtained from patients with cardiovascular pathology following COVID-19, supports the hypothesis that molecular mimicry contributes to the initiation or exacerbation of atherosclerotic cardiovascular disease in the post-COVID period.
A substantial comorbidity between atherosclerotic cardiovascular diseases and coronavirus infections has been consistently reported [1, 2]. The underlying mechanisms of this association remain the subject of active investigation and scientific debate.
Coronaviruses are a large family of viruses that cause respiratory diseases of varying severity, from the common cold to lethal pneumonia. These viruses infect both humans and animals and can induce diseases affecting the respiratory, enteric, hepatobiliary, neuroendocrine, and other physiological systems [3, 4]. To date, seven human coronaviruses have been identified, including the seasonal strains human coronavirus 229E (HCoV-229E), human coronavirus NL63 (HCoV-NL63), human coronavirus OC43 (HCoV-OC43), and human coronavirus HKU1 (HCoV-HKU1), as well as the highly pathogenic viruses severe acute respiratory syndrome coronavirus 1 (SARS-CoV-1), Middle East respiratory syndrome-related coronavirus (MERS-CoV), and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2).
A typical coronavirus virion contains several structurally and functionally significant antigens, including the spike protein (SP), envelope protein (EP), membrane protein (MP), and nucleocapsid protein (NP) [5, 6]. The SP of SARS-CoV-2 and other human coronaviruses plays a key role in mediating viral entry into host cells [7] and represents one of the principal immunodominant viral antigens [8]. It is also a central component of SARS-CoV-2 vaccines. The EP, MP, and NP proteins are likewise mechanistically important and immunogenic molecular targets.
In response to SARS-CoV-2 infection or vaccination, an increase in the intensity of autoimmune processes may occasionally occur. This phenomenon appears to arise in individuals whose immune systems are robust yet somewhat “short-sighted”, relying on mimicking peptides during the immune response. Accordingly, identifying molecular mimicry between coronavirus proteins and human autoantigens may contribute to a deeper understanding of disease pathogenesis, facilitate the development of improved therapeutic approaches, and enhance vaccine safety.
In recent years, a growing body of evidence has documented the promotion, exacerbation, or complication of atherosclerotic cardiovascular disease and/or vasculitides in individuals who have experienced coronavirus disease 2019 (COVID-19) [9–12].
Atherosclerosis may be broadly defined as a chronic, progressive inflammatory condition affecting large and medium-sized elastic and muscular-elastic arteries. It is characterized by a proliferative response of the vascular wall and blood cells to atherogenic lipoproteins, resulting in the formation of atheromas [13–15].
The diversity of existing atherogenesis theories reflects the multifactorial nature of the condition. Among the most influential theories are the response-to-injury hypothesis, the endothelial dysfunction theory, the monoclonal theory, and several infectious theories implicating viruses, Mycoplasma, or Chlamydia. A mosaic of genetic, endocrine, and autoimmune factors is likewise implicated, as these represent critical predispositions and mechanistic contributors to atherogenesis. In addition to classical concepts, an emerging microangiopathic paradigm proposes that atherogenesis may be initiated by vasculitis or ischemia of the vasa vasorum, leading to impaired blood supply and drainage of major arteries [15].
Regarding the proposed mechanisms linking COVID-19 to alterations in the cardiovascular system, several key pathways have been highlighted: the direct cytopathic effect of SARS-CoV-2 on cardiomyocytes, endothelial dysfunction and coagulopathy, an imbalance between myocardial oxygen supply and demand due to compensatory cardiac overstrain in hypoxic conditions, excessive systemic activity of inflammatory mediators, and dysregulation of the renin-angiotensin-aldosterone system [2]. However, the observation that hyperlipidemia in immunodeficient nude mice does not induce atherosclerosis [16] underscores the importance of autoimmune components in atherogenesis [17, 18]. The presumption: “Everything is autoimmune and infectious until proven otherwise”, once coined by Shoenfeld et al. [19], is still valid. The principal concepts of autoimmunity that implicate viral agents as causal factors include epitope spreading [20], cryptic epitope exposure [21], bystander activation [22], and molecular mimicry [23, 24]. Molecular mimicry is defined as the possibility that sequence similarities between foreign and self-peptides may lead to cross-activation of autoreactive T or B cells by pathogen-derived peptides [25].
This phenomenon has been described as “the most suspicious culprit incriminated in the pathophysiology of autoimmunity in the time of COVID-19” [26].
Previously, we demonstrated peptide sharing between immunogenic epitopes of the SARS-CoV-2 SP and several autoantigens of human endocrine cells, accompanied by pathomorphological evidence of autoimmune inflammation in the corresponding endocrine glands in victims of COVID-19 [27, 28].
In the present study, we report new data indicating molecular mimicry between human coronavirus antigens and proteins implicated in atherogenesis and vasculitides. In our view, these findings contribute to a better understanding of the autoimmune component underlying the comorbidity between atherosclerotic cardiovascular complications and coronavirus infection.
The pentapeptide, consisting of a sequence of five amino acids, represents the minimal antigenic determinant capable of inducing highly specific antibodies and mediating defined immune interactions [29, 30]. Therefore, pentapeptides were used as sequence probes. Mimicking pentapeptides were identified using bioinformatic approaches.
Based on published data, we selected 30 human proteins that are either essential participants in atherogenesis or involved in the pathogenesis of atherosclerotic aneurysms and autoimmune arteritides/vasculitides affecting large arteries or the microcirculatory bed. These included: apolipoprotein A-1 (ApoA-1), ApoB-100, ApoC-2, ApoA-3, ApoA(a), oxidized low-density lipoprotein receptor 1, 3-hydroxy-3-methylglutaryl-coenzyme A reductase, proproteinconvertase subtilisin/kexin 9, interstitial collagenase, 72 kDa type IV collagenase, matrix metalloproteinase-9 (MMP9), myeloblastin, myeloperoxidase, 14-3-3 protein sigma, 14-3-3 protein epsilon, 14-3-3 protein theta, 14-3-3 protein beta/alpha, 14-3-3 protein zeta/delta, spermatogenesis-associated protein 7, dihydropteridine reductase, 4-trimethylaminobutyraldehyde dehydrogenase, salivary acidic proline-rich phosphoprotein 2, endothelial protein C receptor, scavenger receptor class B member 1, trimethylaminobutyraldehyde dehydrogenase, carbonic anhydrase 1, microfibril-associated glycoprotein 3, tenascin, migration inhibitory factor-related protein 8 and migration inhibitory factor-related protein 14 [31–44].
Among human coronavirus antigens, we selected the most mechanistically relevant immunogenic proteins: SP, EP, MP, and NP.
All amino acid sequences of human proteins involved in atherogenesis/vasculitides and coronavirus antigens were retrieved from the UniProt database (https://www.uniprot.org/) [45]; corresponding identification numbers are listed in supplementary material (Tables S1–S4). To identify shared minimal immune determinants between human autoantigens and coronavirus antigens, we employed our custom “Alignmentaj” program (available at: https://github.com/muslimb/MyProekt1). The program splits viral protein sequences into pentapeptides (e.g., MFVFL, FVFLV, VFLVL, FLVLL, etc.), and aligns them with the sequences of human proteins implicated in atherogenesis/vasculitides. A detailed description of the algorithm has been provided previously [46].
The immunological relevance of peptide matches was assessed by searching the Immune Epitope Database (IEDB) for immunoreactive epitopes of human coronaviruses containing the shared pentapeptides [47].
Given the recognized involvement of allergic mechanisms [48] and specific cytokines [49] in atherogenesis, we evaluated the allergenic and toxic potential of the mimicking pentapeptides using AllerTOP v2.1 (https://www.ddg-pharmfac.net/AllerTOP/) [50] and ToxinPred tools (https://webs.iiitd.edu.in/raghava/toxinpred/) [51]. All mimicking pentapeptides were also assessed for their IL-10 and IFN-γ induction potential using the IL-10Pred (https://webs.iiitd.edu.in/raghava/il10pred/) [52] and IFNepitope (https://webs.iiitd.edu.in/raghava/ifnepitope/) servers [53].
Based on the bioinformatics findings, enzyme-linked immunosorbent assay (ELISA) was performed in patients with cardiovascular disease following COVID-19. Levels of antibodies against the ApoB-100 (anti-ApoB) (BlueGene, Shanghai, China) and Proteinase 3 (anti-PR3) (Euroimmun, Lübeck, Germany) were measured in serum samples from 26 patients.
To compare the bioinformatic findings with pathomorphological changes in the vessels of persons who died from exacerbation of atherosclerotic cardiovascular disease after previously experiencing COVID-19, a study of autopsy material of the aorta, carotid, and coronary arteries from 6 archive cases was carried out. In six of the studied cases of atherothrombosis, COVID-19 disease, proceeded prior to fatal cardiovascular event, was diagnosed and confirmed by isolating SARS-CoV-2 nucleotide sequences from biological material using the PCR method 2–6 months before death; none of the deceased persons had been vaccinated. At the time of atherothrombotic complications, they all recuperated from COVID-19 with negative tests for SARS-CoV-2. Fragments of vessels for histological examination were fixed with a 10% solution of normal formalin for 72 hours. After fixation, tissue samples were dehydrated using isopropanol, imbibed in paraffin, and embedded in paraffin using standard techniques. Tissue sections 4 µm thick were prepared from paraffin blocks. Histological sections were stained with hematoxylin and eosin, with picrofuchsin by the van Gieson method, and with alcian blue. To detect the expression of CD3 (DASO, clone CD3/Rb a Hu CD3), CD4 (DACO, clone 4B12), CD8 (DACO, clone C8/144B), CD20 (DASO, clone L26), and CD45 (DACO, clone 2B11 + PD7/26), tissue pieces 4 μm thick were prepared from paraffin blocks, placed on treated polylysine glass slides, and immunohistochemical examination was performed. Microscopy was performed with Leica ICC50 W (Leica Microsystems GmbH, Schweiz). The research was approved by the Local Ethical Committee of the L.G. Sokolov North-Western District Scientific and Clinical Centre, approval number 7 dated Dec 08, 2022.
In our study, we investigated molecular mimicry between human coronavirus antigens and human autoantigens related to atherogenesis. The results revealed that the spike (S) protein exhibited the highest degree of pentapeptide sharing with human autoantigens, identifying a total of 93 mimicking peptides, predominantly associated with ApoB-100. The other viral proteins demonstrated progressively fewer shared peptides: the envelope (E) protein had 6 mimicking peptides, primarily linked to ApoB-100; the membrane (M) protein showed 13 mimicking peptides, with a notable association to ApoA-1; and the nucleocapsid (N) protein contained 32 mimicking peptides, mainly corresponding to MMP2. The results of the peptide-matching analysis between the selected human and coronavirus proteins are presented in Tables 1, 2, 3, and 4. Identification numbers (ID numbers) of human coronavirus-derived epitopes containing peptide sequences common to human pentapeptides are shown in square brackets in the supplementary material (Tables S1–S4).
Shared pentapeptides of human proteins and spike proteins of human coronaviruses.
| Key autoantigens | Viruses | Pentapeptide sequences | Total shared pentapeptides |
|---|---|---|---|
| APOA-1 | SARS-CoV-2 | SALGK | 2 |
| HCoV-NL63 | ALEDL | ||
| APOB | SARS-CoV-2 | LFLPF, KLNDL*, SSTAS, ALGKL, DVDLG | 31 |
| SARS-CoV | LFLPF, FKHLR*, KLNDL*, ALGKL, DVDLG | ||
| MERS-CoV | STSAT*, LPVYD*, EVPQL*, SAALS, AALSA, SSFYA | ||
| HCoV-HKU1 | NLKCS*, ESQIS*, LSIQN* | ||
| HCoV-OC43 | KGSVL*, LPDFK*, PDFKE | ||
| HCoV-NL63 | KFSRN, SNASS, PLRLT, SSSGV, LNGNT*, KTLQE | ||
| HCoV-229E | IANII, KTLQE, TLQEL | ||
| APOC2 | SARS-CoV-2 | FLVLL | 2 |
| HCoV-HKU1 | FLVLL | ||
| LPA | HCoV-HKU1 | HRRIP* | 4 |
| HCoV-NL63 | ASSSF, SSSFD, SSFDC* | ||
| HMGCR | MERS-CoV | QLVNA, ALSKL* | 3 |
| HCoV-HKU1 | SLALL | ||
| PCSK9 | SARS-CoV-2 | VLLPL | 5 |
| MERS-CoV | SIQSD*, EDGDY*, FAQSI | ||
| HCoV-OC43 | KSQLV* | ||
| MMP1 | HCoV-OC43 | VTPLT*, LQKAN* | 5 |
| HCoV-NL63 | IQAIY* | ||
| HCoV-229E | YSSFG, IQAIY* | ||
| MMP2 | MERS-CoV | SDGKM* | 2 |
| HCoV-229E | VKFGS | ||
| MMP9 | SARS-CoV | DGKPC | 3 |
| HCoV-OC43 | VTPLT* | ||
| HCoV-229E | SRSEL | ||
| PRTN3 | SARS-CoV-2 | DSFVI*, RAAEI | 3 |
| SARS-CoV | RAAEI | ||
| MPO | SARS-CoV-2 | IVRFP, DQLTP* | 4 |
| SARS-CoV | DQLTP* | ||
| MERS-CoV | SNITI* | ||
| SFN | MERS-CoV | GDYYR, SARSA | 2 |
| YWHAQ | HCoV-OC43 | LIANA | 1 |
| YWHAZ | MERS-CoV | GDYYR | 1 |
| SPATA7 | SARS-CoV-2 | SFIED | 8 |
| SARS-CoV | AVDCS*, SFIED | ||
| MERS-CoV | SVPVS*, VPVSV*, VSTSI* | ||
| HCoV-HKU1 | AVDCS* | ||
| HCoV-229E | GLNTS* | ||
| QDPR | HCoV-NL63 | QSLAG* | 1 |
| SCARB1 | SARS-CoV-2 | GQTGK, QTGKI | 2 |
| ALDH9A1 | HCoV-HKU1 | INNYN | 3 |
| HCoV-OC43 | LGVCV*, KLKDG | ||
| CA1 | MERS-CoV | SKADG | 2 |
| HCoV-HKU1 | LAVIG | ||
| TNC | SARS-CoV-2 | SFSTF | 9 |
| SARS-CoV | TGIGY, ICHEG* | ||
| HCoV-OC43 | TAFAV, LLSRL* | ||
| HCoV-NL63 | DALRL | ||
| HCoV-229E | APEGL*, GLCVD*, LCVDG | ||
| Total | 93 | ||
*: allergen pentapeptide; HCoV-HKU1: human coronavirus HKU1; HCoV-NL63: human coronavirus NL63; HCoV-OC43: human coronavirus OC43; MERS-CoV: Middle East respiratory syndrome-related coronavirus.
Shared pentapeptides of human proteins and envelope proteins of human coronaviruses.
| Key autoantigens | Viruses | Pentapeptide sequences | Total shared pentapeptides |
|---|---|---|---|
| APOB | SARS-CoV-2 | NVSLV | 3 |
| SARS-CoV | NVSLV | ||
| HCoV-HKU1 | QLYKS | ||
| PCSK9 | HCoV-OC43 | KPPVL | 1 |
| MMP9 | SARS-CoV | GVPDL | 2 |
| HCoV-HKU1 | LVLVL | ||
| Total | 6 | ||
*: allergen pentapeptide; HCoV-HKU1: human coronavirus HKU1; HCoV-OC43: human coronavirus OC43.
Shared pentapeptides of human proteins and membrane proteins of human coronaviruses.
| Key autoantigens | Viruses | Pentapeptide sequences | Total shared pentapeptides |
|---|---|---|---|
| APOA-1 | SARS-CoV-2 | ELKKL* | 6 |
| HCoV-HKU1 | TVAKV*, VAKVQ | ||
| HCoV-OC43 | LTIIL, FTIVA, TVAKV* | ||
| HMGCR | HCoV-OC43 | IILTI | 1 |
| PCSK9 | HCoV-NL63 | LVLAL | 2 |
| HCoV-229E | LVLAL | ||
| MMP1 | SARS-CoV | GHSLG | 1 |
| TNC | HCoV-OC43 | WTADE | 1 |
| MRP-8 | SARS-CoV-2 | LKKLL, KKLLE | 2 |
| Total | 13 | ||
*: allergen pentapeptide; HCoV-229E: human coronavirus 229E; HCoV-HKU1: human coronavirus HKU1; HCoV-NL63: human coronavirus NL63; HCoV-OC43: human coronavirus OC43.
Shared pentapeptides of human proteins and nucleocapsid proteins of human coronaviruses.
| Key autoantigens | Viruses | Pentapeptide sequences | Total shared pentapeptides |
|---|---|---|---|
| APOB | SARS-CoV-2 | ALLLL, DDFSK* | 5 |
| SARS-CoV | ALLLL | ||
| HCoV-HKU1 | LKENL | ||
| HCoV-229E | AKFRE* | ||
| APOC2 | HCoV-229E | STAAM | 1 |
| HMGCR | SARS-CoV-2 | LALLL | 2 |
| SARS-CoV | LALLL | ||
| MMP1 | SARS-CoV-2 | KHIDA | 2 |
| SARS-CoV | KHIDA | ||
| MMP2 | SARS-CoV-2 | LGTGP* | 8 |
| SARS-CoV | LGTGP* | ||
| MERS-CoV | FAPGT* | ||
| HCoV-HKU1 | LGTGP* | ||
| HCoV-OC43 | LGTGP* | ||
| HCoV-NL63 | LGTGP* | ||
| HCoV-229E | LGTGP*, SQDDI | ||
| MMP9 | SARS-CoV-2 | ALLLL | 3 |
| SARS-CoV | ALLLL | ||
| HCoV-229E | RGESK | ||
| PRTN3 | SARS-CoV-2 | LALLL | 2 |
| SARS-CoV | LALLL | ||
| MPO | SARS-CoV | SLPYG | 2 |
| HCoV-HKU1 | DFTPE* | ||
| PRH2 | SARS-CoV-2 | GQQQQ, QQQQG* | 4 |
| SARS-CoV | GQQQQ, QQQQG* | ||
| MFAP3 | HCoV-OC43 | LELAK*, ELAKV | 2 |
| TNC | HCoV-229E | SGNTV* | 1 |
| Total | 32 | ||
*: allergen pentapeptide; HCoV-229E: human coronavirus 229E; HCoV-HKU1: human coronavirus HKU1; HCoV-NL63: human coronavirus NL63; HCoV-OC43: human coronavirus OC43; MERS-CoV: Middle East respiratory syndrome-related coronavirus.
The SP of human coronaviruses presented in Table 1 has the largest number of pentapeptides shared with ApoB-100, a structural element of the particles belonging to atherogenic classes of lipoproteins [54, 55]. Tenascin, which is involved in the pathogenesis of aortic aneurysms and was suspected as a target of autoantibodies in aneurysmal disease [40], also has plenty of shared pentapeptides with the SP of human coronaviruses. In addition, myeloperoxidase shows 4 shared pentapeptides. This enzyme oxidizes lipoproteins and makes them more atherogenic [42]. Myeloperoxidase is also a marker antigen of granulomatous vasculitides [56, 57]. Moreover, a study has been published showing that these vasculitides increase the formation of auto-antibody-containing immune complexes as well as modified oxidized lipoproteins, thus accelerating atherogenesis [58]. Although ApoA-1 showed just a single shared pentapeptide with SARS-CoV-2, the increased autoimmunity towards it was registered after COVID-19 [59, 60] and also after vaccination against SARS-CoV-2 [61].
The EPs of human coronaviruses display far fewer shared pentapeptides with human proteins involved in atherogenesis, compared to other viral proteins checked, although several shared pentapeptide sequences have been identified (Table 2). At the same time, among human autoantigens, again three targets directly related to atherosclerosis and its sequelae were identified: ApoB-100, proprotein convertase subtilisin/kexin 9, and MMP9.
Unlike viral SP, the MP of human coronaviruses has far fewer pentapeptides shared with autoantigens (Table 3). The ApoA-1 has the greatest amount of shared pentapeptides with the MP of human coronaviruses. Proprotein convertase subtilisin/kexin, known as the target of treatment by medically used monoclonal antibodies reducing hyperlipoproteinemia [62], and having shared pentapeptides with SP of highly pathogenic coronaviruses, also shows shared pentapeptides with MP of seasonal low-pathogenic ones. Interestingly, spontaneously occurring polyclonal autoantibodies to this enzyme in humans are conversely associated with increased mortality, including atherothrombotic mortality, at least in individuals with diabetes mellitus [63].
The NP of highly pathogenic SARS-CoV-1 and SARS-CoV-2 both display much greater sharing of pentapeptides with investigated autoantigens compared to NP of the seasonal coronaviruses (Table 4).
The autoantigens that have the greatest pentapeptide similarity with viral NPs are MMPs, MMP2 and MMP9, which play a significant role in the formation and existence of atherosclerotic aortic aneurysms [64]. Autoimmunity to these enzymes in coronavirus infection might have acted protectively rather than pathogenically, this being antiatherogenic. In experiments, monoclonal antibodies to these collagenases inhibited the transformation of smooth myocytes in atheromas into foam cells and reduced the atherogenicity of oxidized lipoproteins [65]. The immunologically achieved suppression of MMP2 and MMP9 activity was protective in experimental abdominal aortic aneurysm formation [44, 66].
The positive predictions of allergenicity of shared mimicking pentapeptides are presented in Table 1, Table 2, Table 3, and Table 4 as asterisk matched. None of the 144 viral shared pentapeptides has the potency to stimulate IL-10 production; on the contrary, all of them were γ-interferonogenic. All 144 shared pentapeptides were devoid of toxicity potential, according to the ToxinPred tool.
In the next step, to confirm the bioinformatic analyses, the ELISA studies were performed to detect autoantibodies against human atherogenesis autoantigens in patients with cardiovascular disease after COVID-19. Antibodies against the ApoB-100 (anti-ApoB) and PR3 (anti-PR3) were measured in 26 patients with cardiovascular disease after COVID-19.
The statistical analysis of anti-ApoB antibodies in patients with cardiovascular disease following COVID-19 reveals a median level of 20.24 ng/mL, indicating that half of the patients exhibit antibody levels below this threshold. The first quartile (Q1) is 0.6, suggesting that 25% of the cohort has minimal or undetectable antibody levels, while the third quartile (Q3) at 45.82 indicates that 75% of patients have levels below this value. The interquartile range (IQR) between Q1 and Q3 highlights substantial variability in antibody responses, with a notable proportion of patients presenting low levels, contrasted by a significant subset with elevated levels. These findings suggest a heterogeneous immune response to ApoB in this population, warranting further investigation into the clinical implications of these varying antibody levels in the context of post-COVID-19 cardiovascular outcomes.
Out of the 26 patients with cardiovascular disease following COVID-19, 4 patients were found to have elevated levels of anti-PR3 antibodies (reference values > 20 RU/mL) (median: 5.45, Q1: 0.53, Q3: 12.9). This indicates a potential association between high anti-PR3 levels and the development of cardiovascular complications in this group of patients post-COVID-19.
Gross examination of the aorta, carotid, and coronary arteries revealed yellow to whitish-yellow plaques, some of which coalesced into larger formations. Some plaques in the aorta had smooth, dense surfaces. Some plaques instead demonstrated hemorrhages or thrombotic deposits. Similar circular plaques were observed in coronary and carotid arteries.
Histological analysis identified plaques of varying types, including:
plaques with thin connective-tissue caps and substantial lipid deposits (unstable);
plaques with thick fibrous caps rich in collagen, with small lipid cores (stable);
plaques exhibiting mechanical disruption, tears, and ruptures of the fibrous cap (complicated).
The adventitia contained clusters of microvessels with thickened walls, indicating established vascularization of the arterial wall. In stable plaques, the vasa vasorum displayed uniform wall thickness and lacked glycosaminoglycan deposits (confirmed by alcian blue staining). In the areas of unstable plaques, the vasa vasorum showed uneven wall thickening due to focal glycosaminoglycan deposition (Figure 1), endothelial degeneration, and mild-to-moderate perivascular lymphocytic infiltration with edema (Figure 2). Immunohistochemistry demonstrated infiltration by CD3+, CD8+, and CD20+ lymphocytes, with CD20+ cells predominating (Figure 3).
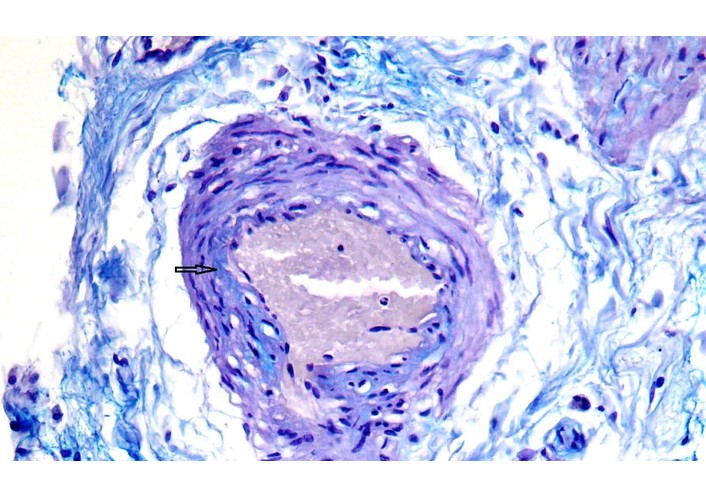
Representative histopathological image from an unstable plaque region of vasa vasorum in the adventitia of the aorta: focal deposits of glycosaminoglycans in the vessel wall (arrow). Аlcian blue stain, ×400.
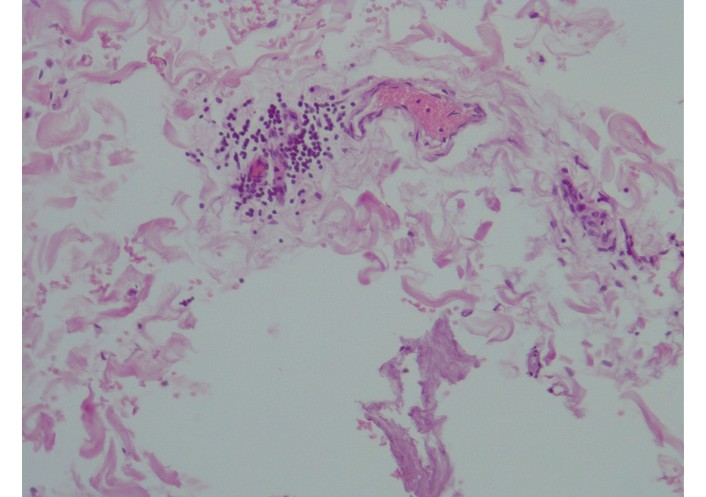
Representative histopathological image from an unstable plaque region of the adventitia of the aorta. Focal lymphocytic infiltrate around vasa vasorum, hematoxylin & eosin stain, ×200.
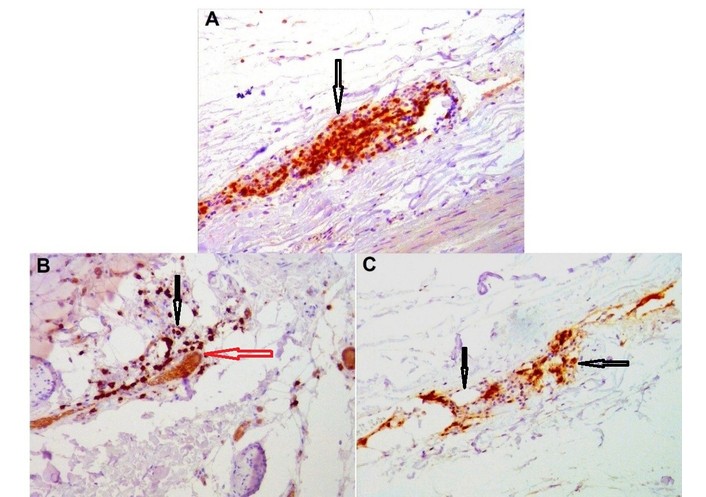
Representative immunohistochemical image from an unstable plaque region of perivascular infiltrate in the adventitia of the coronary artery. A. CD3 (arrows), ×100; В. CD8 (red arrow—vasa vasorum, black arrows—CD8+ lymphocytes), ×200; С. CD20 (black arrows—CD20+ lymphocytes), ×200.
Several general considerations regarding antigenic mimicry merit emphasis. Beyond the direct pathological effects of coronavirus infection on cardiovascular tissues, indirect immune-mediated mechanisms are plausible. Previously, we documented molecular mimicry between SARS-CoV-2 antigens and host autoantigens implicated in autoimmune endocrinopathies [28], with accompanying lymphocytic inflammation of multiple endocrine glands in fatal COVID-19 cases. Such lymphoid infiltration reflects cell-mediated immunopathology.
In the context of post-COVID cardiovascular disease, endocrine involvement—particularly thyroid dysfunction—may be relevant. Hypothyroidism is strongly associated with atherogenesis, owing to alterations in lipid metabolism [67] and the frequent presence of secondary arterial hypertension [68], both of which are established risk factors of atherosclerosis. Autoantibody formation after infection is widely attributed to molecular mimicry between pathogen antigens and self-antigens [69].
Our bioinformatic and serological findings support a potential role for mimicry in atherogenesis. The study demonstrates sequence homology and correlational findings, such as the presence of antibodies and inflammation; however, it lacks functional validation to confirm that the shared peptides can trigger cross-reactive immune responses leading to functional pathology. Therefore, the Discussion emphasizes that the proposed mechanism of molecular mimicry is supported by circumstantial evidence but not definitively proven.
The human immune system exhibits differential responses to various microorganisms, primarily attributable to human leukocyte antigen (HLA) haplotypes. It is posited that the development of autoimmunity is associated with human HLA. When shared peptides bind to human HLA and are presented on the surface of antigen-presenting cells, the potential for autoimmune reactions may arise within the human body.
Antigenic mimicry constitutes a prerequisite rather than a guarantee of post-infectious autoimmunity. Antigen-presenting cells may present different protein fragments depending on individual HLA haplotypes, as demonstrated for infectious triggers of autoimmune encephalitides [70]. Nevertheless, in post-COVID atherothrombosis, we observed productive inflammation with lymphoid infiltrates surrounding the vasa vasorum in regions of unstable plaques. Such changes are characteristic of autoimmune vascular inflammation, as described in various vasculitides and in atherosclerotic aneurysms [71, 72]. Our findings of both T- and B-cell infiltration align with the concept that molecular mimicry may mislead both T-cell receptors and B-cell immunoglobulins. Involvement of the vasa vasorum (“vasa-vasoritis”) is typical of infection-triggered aortitis [73] and has long been considered a contributing factor in atherogenesis [74]. Cases of COVID-19-associated arteritis, aortitis, and systemic vasculitides have also been reported [75].
All shared pentapeptides identified in this study were predicted to induce IFN-γ, but not IL-10, and 45 shared pentapeptides across various coronaviruses were predicted to be potentially allergenic.
Innate immunity—particularly natural killer (NK) cell activation in response to reduced HLA class I expression on metabolically impaired or virus-infected cells—is a key early component of antiviral defense. Activated NK cells release perforin, granzyme B, and IFN-γ [76, 77]. IFN-γ upregulates transcription of numerous immune-related genes [78] and enhances antigen presentation by shifting proteasome activity toward immunoproteasomes [79]. Excessive immune stimulation is a hallmark of autoimmune pathology, including autoimmune aspects of atherogenesis. The predicted allergenicity of 45 shared pentapeptides is noteworthy, as IgE-mediated allergies represent hyperreactive immune states and have been linked to an increased risk of atherosclerotic cardiovascular disease [80] as well as to COVID-19 pathogenesis [81].
In the absence of control groups, the proposed causal link between coronavirus infection, molecular mimicry, and the observed pathology remains speculative. While our observations suggest an association, we cannot confirm causation. Therefore, our conclusions should be approached with caution, highlighting the need for further research with appropriate controls to validate these findings and elucidate the underlying mechanisms.
In conclusion, despite extensive research, no unified theory of atherogenesis currently exists. Infectious and autoimmune mechanisms remain central components of its etiopathogenesis. As Khan et al. [82] (2024) observed, “atherosclerosis triggers the breakdown of tolerance to self-components” [82]. Bioinformatic analysis represents an important step toward elucidating the potential molecular mimicry between exogenous pathogens and human proteins. Our findings of sequence similarity between human coronavirus proteins and autoantigens associated with atherogenesis and arteritides—supported by the corresponding pathomorphological and serological evidence—suggest that COVID-19 may represent a novel factor contributing to the immunopathological mechanisms underlying atherosclerotic cardiovascular disease.
The study provides hypothesis-generating evidence for a potential role of COVID-19 via molecular mimicry in atherosclerotic disease, which requires validation through controlled, functional studies.
It is essential to emphasize the importance of further morphometric research to gain a deeper understanding of the obtained results on lymphocytic infiltration around the vasa vasorum. We also recommend conducting serological studies with case controls to validate our findings, as well as utilizing in vitro analysis of T-cell and B-cell cross-reactivity with peptide-stimulated T-cell proliferation assays or immunization models. Additionally, it would be beneficial to consider conducting studies in animal models for a functional test of the mimicry hypothesis, which could provide further insights into the mechanisms underlying our observations.
In our view, further laboratory experiments verifying antigenic cross-talk and its functional consequences could be organized in the same way as previously used in our study of the molecular mimicry of coronavirus proteins with antigens involved in spermatogenesis [83]. Proteins could be synthesized from mimicking peptides, followed by testing for the presence of autoantibodies to them in patients and controls, and then the immunogenicity and pathogenicity studies can be performed by immunizing mice with selected peptides and testing the antigenicity of induced antibodies. The influence of such immunization on the course and severity of experimental atherosclerosis can also be checked.
The study limitations are:
One of the primary limitations of this study is the relatively small sample size. A limited number of participants may affect the generalizability of the findings and reduce the statistical power of the analyses. Future research should aim to include a larger and more diverse sample to enhance the robustness of the results and ensure that they are representative of the broader population.
A critical limitation of this study is the absence of control groups for ELISA analysis of antibodies against the ApoB-100, which hinders our ability to establish definitive causal relationships between coronavirus infection, molecular mimicry, and the observed pathology.
ApoB-100: apolipoprotein B-100
COVID-19: coronavirus disease 2019
ELISA: enzyme-linked immunosorbent assay
EP: envelope protein
HCoV-229E: human coronavirus 229E
HCoV-HKU1: human coronavirus HKU1
HCoV-NL63: human coronavirus NL63
HCoV-OC43: human coronavirus OC43
HLA: human leukocyte antigen
IFN: interferon
IL-10: interleukin-10
MERS-CoV: Middle East respiratory syndrome-related coronavirus
MMP9: matrix metalloproteinase-9
MP: membrane protein
NK: natural killer
NP: nucleocapsid protein
PR3: Proteinase 3
SARS-CoV-1: severe acute respiratory syndrome coronavirus 1
SARS-CoV-2: severe acute respiratory syndrome coronavirus 2
SP: spike protein
The supplementary tables for this article are available at: https://www.explorationpub.com/uploads/Article/file/1003257_sup_1.pdf.
LPC: Conceptualization, Funding acquisition, Methodology, Project administration, Supervision, Writing—review & editing. VSG: Methodology, Validation. MVM: Methodology. TAN: Methodology, Writing—original draft. VAS: Methodology. PAS: Methodology. VJU: Data curation, Writing—original draft, Writing—review & editing. TVF: Data curation. MGN: Conceptualization, Formal analysis, Investigation, Methodology, Resources, Software, Visualization, Writing—original draft. All authors read and approved the submitted version.
The authors declare that they have no conflicts of interest.
This study complies with the Declaration of Helsinki (2013), and it was approved by the Local Ethical Committee of the L.G. Sokolov North-Western District Scientific and Clinical Centre, approval number 7 dated Dec 08, 2022.
Informed consent to participate in the study was obtained from all participants.
Not applicable.
All relevant data are available from the corresponding author on request (muslimbek.normatov@gmail.com)
This work was supported by the grant of the Government of the Russian Federation for the state support of scientific research carried out under the supervision of leading scientists, agreement NO. 075-15-2022-1110 dated June 30, 2022. Scientific research was performed at the Laboratory of Microangiopathic Mechanisms of Atherogenesis of St. Petersburg State University. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 396
Download: 23
Times Cited: 0
