 Original Article
Original Article

Affiliation:
1Department of Emergency Medicine, Saveetha Medical College and Hospital, Saveetha Institute of Medical and Technical Sciences (SIMATS), Saveetha University, Chennai 602105, Tamil Nadu, India
ORCID: https://orcid.org/0000-0001-6517-4536
Affiliation:
2Post Graduate and Research Department of Biotechnology & Microbiology, National College (Autonomous), Tiruchirapalli 620001, India
Affiliation:
2Post Graduate and Research Department of Biotechnology & Microbiology, National College (Autonomous), Tiruchirapalli 620001, India
Affiliation:
3Department of Biotechnology, School of Integrative Biology, Central University of Tamil Nadu, Thiruvarur 610005, India
ORCID: https://orcid.org/0000-0001-8973-3507
Affiliation:
4Department of Molecular Biology, Beykoz Institute of Life Sciences & Biotechnology (BILSAB), Bezmialem Vakif University, Istanbul 34820, Türkiye
ORCID: https://orcid.org/0009-0001-8128-3991
Affiliation:
2Post Graduate and Research Department of Biotechnology & Microbiology, National College (Autonomous), Tiruchirapalli 620001, India
Email: mohamedjaabir@nct.ac.in
ORCID: https://orcid.org/0000-0002-4880-9939
Explor Foods Foodomics. 2026;4:1010177 DOI: https://doi.org/10.37349/eff.2026.1010177
Received: January 20, 2026 Accepted: June 17, 2026 Published: July 09, 2026
Academic Editor: Maria Tsimidou, Aristotle University of Thessaloniki, Greece
Aim: Maternal nutritional insufficiency during pregnancy is a major public health concern associated with adverse neurodevelopmental outcomes in offspring. This study investigated the effects of global maternal nutrient restriction before and/or during gestation on cognitive and behavioral performance in rat offspring and evaluated the potential for recovery following postnatal dietary normalization.
Methods: Female Wistar rats were assigned to either a control diet (AIN-93G) or a 50% global nutrient-restricted diet. Nutrient-restricted groups received dietary restriction before pregnancy alone (RD-P) or before and throughout gestation (RD-W), followed by a standard diet after birth or weaning. Offspring were assessed at postnatal days 35–40 using the Y-maze spontaneous alternation test for working memory, the Open Field Test for locomotor activity, and the Elevated Plus Maze (EPM) for anxiety-like behavior. Data were analyzed using one-way ANOVA with Dunnett’s post hoc test (p < 0.05).
Results: Maternal nutrient restriction did not significantly affect Y-maze spontaneous alternation performance (p > 0.05), indicating preserved working memory. However, offspring from restricted groups exhibited significantly greater time spent in EPM open arms (p < 0.001), suggesting reduced anxiety-like behavior, with the strongest effect observed in the RD-W group. Increased locomotor activity was also detected in nutrient-restricted offspring during the Open Field Test (p = 0.0118). Gestational maternal food intake showed weak negative correlations with EPM open-arm duration (r = −0.34, p = 0.06) and locomotor activity (r = −0.28, p = 0.12).
Conclusions: Maternal global nutrient restriction induces persistent, domain-specific behavioral alterations in offspring. While postnatal dietary normalization may partially ameliorate deficits in activity-related behaviors, reduced anxiety-like behavior persists into adolescence. These findings emphasize the critical role of adequate maternal nutrition in shaping offspring neurodevelopment and behavioral health, highlighting the importance of nutritional interventions during the preconceptional and gestational periods.
Cognitive management, impulse control, learning ability, and emotional modulation are all critical cognitive and behavioral processes for normal neurodevelopment. Changes in these processes throughout early childhood can cause long-term changes in brain function and behavior, altering academic success, social relationships, and overall quality of life across the lifetime [1]. Attention deficits are frequently characterized by poor sustained concentration and executive organization, whereas hyperactivity shows excessive motor drive and difficulties maintaining behavioral constraint. Impaired impulse control is also seen as fast, poorly controlled reactions and less executive monitoring [2]. Neurodevelopmental outcomes are the result of a complex interaction between genetic predisposition and environmental effects, particularly during the prenatal and early postnatal periods, when the brain grows rapidly and circuits emerge [3]. The integrity of dopaminergic and noradrenergic signaling pathways, which are essential for executive function, emotional control, and adaptive behavior, is a major determinant of cognitive and behavioral regulation at the neurobiological level [4]. Cognitive and behavioral vulnerabilities can remain if these neurotransmitter systems are disturbed throughout development [5]. Maternal nutrition throughout pregnancy has become one of the most important environmental factors influencing the development of the offspring’s brain. Adequate micronutrient availability is required for neuronal proliferation, myelination, synaptogenesis, and neurotransmitter production [6]. Micronutrients including iron, zinc, iodine, folate, vitamin B12, and omega-3 fatty acids are especially vital during critical stages of fetal brain development, when deficiencies can permanently affect neuronal architecture and plasticity [7]. For instance, a lack of iron during pregnancy has been linked to long-term cognitive and attention problems, altered dopamine metabolism, and decreased myelination [8]. Similarly, it has been demonstrated that shortages in omega-3 polyunsaturated fatty acids affect cortical development, synaptic transmission, and neuronal membrane integrity, which results in deficits in cognition and emotional control [9]. Deficits in zinc and iodine are also associated with decreased neurogenesis, altered neurotransmission, and worse cognitive function in children [10].
Approximately 30 to 40 percent of pregnant women worldwide suffer from many micronutrient deficiencies. In low-resource contexts, macronutrient undernutrition affects around 20% of pregnancies, whereas anemia affects approximately 45–50% of women in India who are of reproductive age [11–13]. Even though the significance of prenatal nutrition is becoming more widely acknowledged, the majority of the literature to date has concentrated on postnatal dietary interventions and childhood nutrition, with relatively few studies looking at the long-term cognitive effects of maternal nutrient deficiencies during pregnancy [14]. This is a major knowledge gap on how early dietary settings affect later-life behavioral regulation and cognitive pathways. Nutritional deficiencies can have long-lasting consequences on brain structure and function throughout the perinatal period, which is a particularly sensitive window [15]. These developmental processes may be studied in controlled experimental settings using preclinical animal models. Maternal nutritional changes and their neurobehavioral effects on offspring may be systematically evaluated thanks to rodent models, which in particular accurately mimic important features of human neurodevelopment and cognitive circuitry [16]. Mechanistic insights into the effects of prenatal nutritional stress on cognitive control, activity levels, emotional response, and executive function are made possible by these models, which enable in-depth examination of molecular, cellular, and behavioral endpoints [17]. The current study examines the long-term behavioral and cognitive effects of maternal nutritional deprivation using a rat model. This work attempts to clarify the mechanisms via which inadequate maternal nutrition affects child brain development and cognitive function by evaluating neurobehavioral outcomes in addition to developmental indices [18]. In order to promote normal neurodevelopment and lessen the burden of cognitive and behavioral deficits across communities, evidence-based dietary regimens, public health policies, and early interventions may be informed by an understanding of these pathways [19].
The Institutional Animal Ethical Committee (IAEC), National College, Tiruchirappalli, India, and the European Communities Council Directive for the care and use of laboratory animals were strictly followed during all animal operations and were authorized prior to the start of the study under CPCSEA registration number 1941/GO/RE/S/17/CPCSEA. We purchased adult Wistar rats from the Indian Institute of Science in Bangalore. The focus of this study was only behavior. No animals were slaughtered or intrusive treatments carried out. Following behavioral evaluations, all rats were treated humanely and kept in the institutional animal facility under normal living circumstances in compliance with the Guide for the Care and Use of Laboratory Animals and CPCSEA.
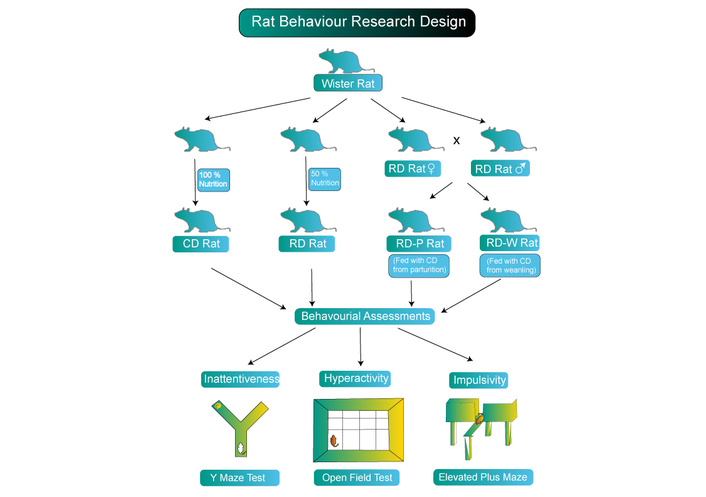
Virgin female Wistar rats (12–14 weeks old, no siblings) were kept under standard circumstances (temperature 22 ± 2°C, humidity 55 ± 5%, 12 h light/dark cycle) with unrestricted access to tap water and standard rat food. After 7 days of acclimatization, males were paired with females periodically until mating was verified by the presence of a vaginal copulation plug (1–4 days later). Females were randomly allocated to four groups (initial n = 14 each):
The CD (Control Diet) followed the usual AIN-93G diet throughout.
RD (Restricted Diet): 50% global nutrition restriction throughout gestation.
RD-P (Limited Diet – Pre-pregnancy): limited diet prior to pregnancy, transitioning to CD postpartum.
RD-W (Limited Diet – Weaning): limited diet prior to and throughout pregnancy, transitioning to CD after weaning (postnatal day 21).
The purpose of the RD-P and RD-W groups was to differentiate between the effects of gestational restriction and preconception on children. Non-pregnant females were eliminated after mating (RD-P: 6 excluded → 8 pregnant; RD-W: 4 excluded → 10 pregnant). For behavioral testing, offspring from pregnant mothers (n = 8–10 pups/group, balanced sex ratio) were utilized.
From gestation day 0 to postnatal day 21 (weaning), pregnant rats were placed on either the restricted diet (RD; 50% of all nutrients, both macro and micronutrients) or the control diet (CD; AIN-93G standard, 100% nutritional needs; Reeves et al., 1993) [20]. Diets were obtained from a certified commercial supplier. The RD-P and RD-W groups were transferred to CD during weaning and after giving birth, respectively. Every week, body weight and food intake were recorded (refer to Results, Table 1).
Maternal food intake (g/day, mean ± SD) and body weight (g, mean ± SD) during gestation and lactation.
| Parameter | CD | RD | RD-P | RD-W |
|---|---|---|---|---|
| Food intake (g/day) | ||||
| Gestation Week 1 | 18.4 ± 1.2 | 9.2 ± 0.8 | 9.0 ± 0.9 | 9.1 ± 0.7 |
| Gestation Week 2 | 19.1 ± 1.4 | 9.6 ± 0.9 | 9.4 ± 1.0 | 9.5 ± 0.8 |
| Gestation Week 3 | 20.3 ± 1.5 | 10.1 ± 1.0 | 19.8 ± 1.3† | 10.0 ± 0.9 |
| Lactation Week 1 | 32.5 ± 2.8 | 16.8 ± 1.7 | 31.9 ± 2.6 | 17.2 ± 1.6 |
| Lactation Week 2 | 38.2 ± 3.1 | 19.4 ± 2.0 | 37.8 ± 3.0 | 19.1 ± 1.9 |
| Lactation Week 3 | 35.6 ± 2.9 | 18.2 ± 1.8 | 35.1 ± 2.7 | 18.0 ± 1.7 |
| Body weight (g) | ||||
| Gestation week 0 | 248 ± 12 | 245 ± 11 | 247 ± 10 | 246 ± 12 |
| Gestation week 1 | 262 ± 14 | 252 ± 12 | 254 ± 11 | 253 ± 13 |
| Gestation week 2 | 285 ± 16 | 258 ± 13 | 260 ± 12 | 259 ± 14 |
| Gestation week 3 | 318 ± 18 | 265 ± 14 | 268 ± 13 | 266 ± 15 |
| Lactation week 1 | 302 ± 15 | 258 ± 13 | 295 ± 14 | 260 ± 14 |
| Lactation week 2 | 288 ± 14 | 252 ± 12 | 282 ± 13 | 254 ± 13 |
| Lactation week 3 | 275 ± 13 | 248 ± 11 | 270 ± 12 | 250 ± 12 |
†: Indicates a statistically significant difference compared to the control group (p < 0.05).
The offspring were evaluated for attention, hyperactivity, and behaviors associated with anxiety or impulsivity between postnatal days 35 and 40 (adolescence/young adulthood) [19–22]. After acclimating to the testing room for one hour, all tests were carried out during the light phase. In order to prevent olfactory signals, the apparatus was cleaned between experiments.
The arms of the Y-maze measured 40 cm in length, 3 cm in bottom width, 13 cm in upper width, and 15 cm in wall height. Rats were positioned in the middle, and video tracking was used to capture their spontaneous alternation for ten minutes.
An arena of 50 cm × 50 cm × 50 cm with a center zone 10 cm from walls was utilized for the Open Field Test (locomotor activity/anxiety-like behavior). Ten minutes of time spent in the center and the total distance travelled were noted.
A plus-shaped maze was employed, with closed arms and 20 cm black walls. The arms were 40 cm long, 10 cm wide, and 50 cm high. Rats began by standing in the middle with their arms spread wide; their time there was noted for five minutes, which is the typical amount of time needed to measure anxiety.
All statistical analyses were performed using GraphPad Prism software (Version 10.2.0; GraphPad Software Inc., San Diego, CA, USA). Data are presented as mean ± standard deviation (SD). Comparisons among experimental groups were conducted using One-way ANOVA and Dunnett’s post-hoc test (control vs. each RD group) to examine the data. The mean ± SD is used to show the results. The threshold for significance was p < 0.05. There are 8–10 pups per batch. Every test is carried out as indicated in the Supplementary materials.
Throughout gestation and lactation, maternal food consumption and body weight were recorded weekly (Table 1). The RD and RD-W groups consumed 50% less food than the CD group during restriction periods (p < 0.001). Following the transfer to a control diet, food intake in RD-P normalized after parturition, and in RD-W after weaning. Compared to CD, restricted groups (RD and RD-W) saw substantially lower body weight growth throughout gestation (p < 0.01), indicating decreased calorie and nutrient consumption. After diet normalization, postnatal weight trajectories in the RD-P and RD-W groups improved to some extent. Pearson correlation studies revealed a modest negative relationship between gestational food intake and offspring Elevated Plus Maze (EPM) open-arm time (r = –0.34, p = 0.06), indicating that decreased maternal nutrition intake during pregnancy may lead to less anxiety-like behavior in children. There were no significant relationships between maternal food/body weight and Y-maze alternation or Open Field distance (p > 0.05) (Table 2).
Pearson correlation coefficients (r) between maternal parameters and offspring behavioral outcomes.
| Maternal parameter | Y-maze alternation (%) (working memory) | EPM open-arm time (s) (anxiety-like behavior) | Open Field distance traveled (cm) (locomotor activity) |
|---|---|---|---|
| Gestational food intake (g/day, avg) | r = 0.12 (p = 0.48) | r = –0.34 (p = 0.06) | r = –0.28 (p = 0.12) |
| Gestational body weight gain (g) | r = 0.09 (p = 0.61) | r = –0.31 (p = 0.08) | r = –0.25 (p = 0.18) |
| Lactational food intake (g/day, avg) | r = 0.08 (p = 0.65) | r = –0.19 (p = 0.32) | r = –0.15 (p = 0.44) |
Correlations calculated across all groups (n ≈ 36–40 pups with maternal data). Negative r values indicate that lower maternal intake/weight gain was associated with more open-arm time (reduced anxiety-like behavior) and increased locomotor activity. No correlations reached statistical significance (all p > 0.05), but trends were observed for gestational parameters and EPM/Open Field outcomes.
No significant differences in alternation scores across groups (one-way ANOVA, F(3,36) = 1.45, p = 0.24). The RD group showed numerically higher scores compared to CD (mean difference = 18.00, 95% CI: –9.47 to 45.47, p = 0.21). RD-P and RD-W groups did not differ from CD (p = 0.61 and p = 0.95, respectively) (Figure 1).
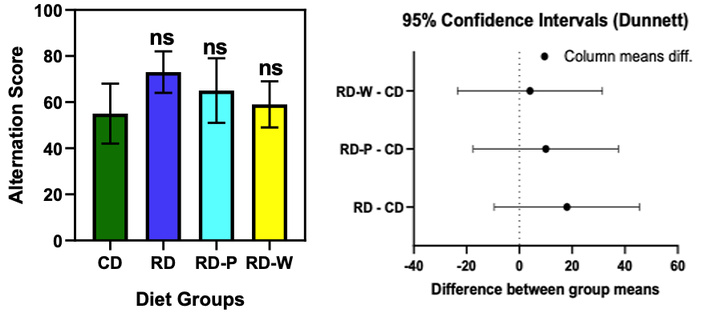
Spontaneous alternation test results comparing control diet (CD), restricted diet (RD), postnatal restricted diet (RD-P), and weaning restricted diet (RD-W) groups. No significant differences in alternation scores were observed between the diet groups, indicating that dietary restriction did not significantly impact cognitive flexibility or inattentiveness. The RD group showed a slightly higher alternation score compared to the CD group, but the differences were not statistically significant (p > 0.05). Statistical notation: ns = not significant (p ≥ 0.05); * = p < 0.05; ** = p < 0.01; *** = p < 0.001; **** = p < 0.0001 vs. CD group according to Dunnett’s multiple-comparison test.
Significant group effect on open-arm time (one-way ANOVA, F(3,36) = 12.34, p < 0.0001). All restricted groups spent more time in open arms compared to CD, indicating reduced anxiety-like behavior: RD (mean difference = 28.5 s, 95% CI: 17.55–39.45, p = 0.0002), RD-P (mean difference = 25.5 s, 95% CI: 14.55–36.45, p = 0.0004), RD-W (mean difference = 51.5 s, 95% CI: 40.55–62.45, p < 0.0001) (Figure 2).
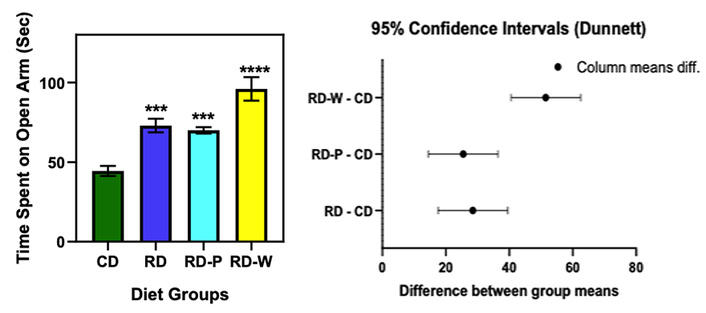
Elevated plus maze (EPM) performance of offspring from different maternal diet groups. Time spent in the open arms was significantly increased in the restricted diet (RD), postnatal restricted diet (RD-P), and weaning restricted diet (RD-W) groups compared with the control diet (CD) group, indicating reduced anxiety-like behavior. The RD-W group exhibited the highest open-arm duration, suggesting the strongest anxiolytic-like effect among the dietary restriction groups. Data are presented as mean ± SD. Statistical analysis was performed using one-way ANOVA followed by Dunnett’s multiple-comparison test against the CD group. Statistical notation: ns = not significant (p ≥ 0.05); * = p < 0.05; ** = p < 0.01; *** = p < 0.001; **** = p < 0.0001 vs. CD group according to Dunnett’s multiple-comparison test.
Significant increase in distance traveled in RD vs. CD (mean difference = 31.00, 95% CI: 7.99–54.01, p = 0.0118). No significant differences in RD-P (p = 0.92) or RD-W (p = 0.9985) (Figure 3).
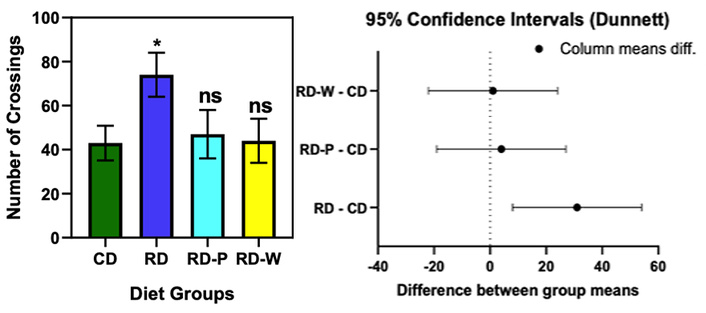
Open Field Test assessment of locomotor activity in offspring from different maternal diet groups. The restricted diet (RD) group showed significantly increased locomotor activity compared with the control diet (CD) group, indicating hyperactivity associated with maternal nutrient restriction. No significant differences were observed in the postnatal restricted diet (RD-P) and weaning restricted diet (RD-W) groups relative to CD, suggesting partial recovery following dietary normalization. Data are presented as mean ± SD. Statistical analysis was performed using one-way ANOVA followed by Dunnett’s multiple-comparison test against the CD group. Statistical notation: ns = not significant (p ≥ 0.05); * = p < 0.05; ** = p < 0.01; *** = p < 0.001; **** = p < 0.0001 vs. CD group according to Dunnett’s multiple-comparison test.
During crucial developmental windows, maternal global food restriction caused domain-specific behavioral changes in rat offspring. There was no discernible difference in working memory across groups as measured by spontaneous alternation in the Y-maze (p > 0.05). This result is consistent with a number of earlier studies that found that moderate gestational nutrient restriction had mild or context-dependent effects on attention and executive function. This could be due to compensatory hippocampus mechanisms or the short duration of restriction in the current model [18, 19]. In keeping with studies that attentional/executive deficits are frequently modest or task-specific in such contexts, the lack of a substantial deficit in alternation performance raises the possibility that working memory circuits may be resilient to mild prenatal undernutrition [20, 21].
On the other hand, all restricted groups showed a substantial increase in time spent in the open arms of the EPM, which demonstrated a reduction in anxiety-like behavior (p < 0.001). The highest impact was shown in RD-W offspring (p < 0.0001). This paradoxical decrease in anxiety-like traits is in line with new research that suggests undernutrition during pregnancy can reduce stress responsiveness and anxiety-related behaviors in rodent models. This may be due to changes in frontostriatal maturation, altered HPA-axis programming, or decreased threat sensitivity [22, 23, 24]. The idea that prenatal food restriction may rewire emotional regulatory circuitry toward less anxiety-like responses is supported by the fact that enhanced open-arm exploration is traditionally viewed as decreased anxiety-like behavior rather than increased impulsivity [25, 26].
Only the RD group showed a significant increase in hyperactivity, as shown by the total distance walked in the Open Field Test (p = 0.0118); following postnatal diet normalization, there were no discernible differences between the RD-P or RD-W groups. This pattern implies that postnatal dietary recovery permits partial flexibility in motor and arousal systems, whereas gestational restriction has a more noticeable impact on locomotor circuits [27, 28]. In keeping with differing developmental windows of sensitivity, the partial mitigation shown in the RD-P and RD-W groups further suggests that locomotor hyperactivity could be more responsive to postnatal intervention than anxiety-like traits [19, 29].
Moderately unfavorable trends were found by Pearson correlation analyses between the open-field distance (r = –0.28, p = 0.12) and EPM open-arm time (r = –0.34, p = 0.06) and gestational maternal food consumption. These correlations imply that whereas postnatal recovery of intake largely normalizes these effects, poorer nutritional availability during pregnancy may lead to decreased anxiety-like behavior and increased locomotor activity in offspring. Working memory may be less responsive to the level of maternal constraint in this paradigm, as seen by the lack of substantial associations with Y-maze performance. Dopaminergic synthesis, myelination, and the formation of the frontostriatal circuitry depend on a number of micronutrients, especially iron, zinc, choline, folate, and vitamin B12 [30, 31]. Iron deficiency causes dysregulation of attention and impulse control by impairing dopamine metabolism and D2 receptor function [32, 33]. While folate and B12 deficits impair methylation pathways and raise homocysteine, which impacts brain plasticity and synaptic maturation, zinc and choline promote neurogenesis and cholinergic transmission [34, 35]. Through energy deficit, changed insulin/IGF-1 signaling, and decreased substrate availability for brain development, macronutrient restriction most likely exacerbated these effects [36, 37]. Although the lack of direct neurochemical (dopamine levels, receptor expression) or neuroanatomical (prefrontal cortex histology) measurements in this study means that such interpretations remain conjectural and need further validation, these pathways offer tenable mechanisms for the observed behavioral profile.
This study has a few drawbacks, including lower final sample numbers owing to non-pregnant exclusions. To identify micronutrient vs calorie effects, no negative controls (for example, a pair-fed isocaloric group) were used. The 21-day gestational restriction model represents moderate exposure but may not depict chronic deficit. No molecular, neurochemical, or histological studies were performed. Behavioral paradigms give indirect assessments of ADHD-like features, but rodent models have intrinsic limitations in properly replicating human neuropsychiatric disorders. Future research should precisely distinguish between macronutrient and micronutrient impacts, target specific nutrients (iron, zinc, choline, B-vitamins), and use sophisticated models (patient-derived organoids, humanized mice, multi-omics) to unravel processes. Longitudinal designs with longer restriction periods, neurochemical assessments (e.g., dopamine levels, receptor expression), and brain histology are encouraged.
This study shows that maternal global nutritional restriction during pregnancy causes long-term behavioral changes in children, including reduced anxiety-like behavior and hyperactivity, which may be reversed partially after birth. These findings emphasize the importance of sufficient maternal nutrition in neurodevelopmental outcomes and encourage early nutritional treatments to reduce cognitive and behavioral vulnerabilities.
CD: control diet
EPM: Elevated Plus Maze
RD: restricted diet
RD-P: postnatal restricted diet
RD-W: weaning restricted diet
The supplementary materials for this article are available at: https://www.explorationpub.com/uploads/Article/file/1010177_sup_1.pdf.
The authors gratefully acknowledge the financial support provided by the University Grants Commission (UGC), New Delhi, Government of India. The authors also thank National College (Autonomous), Tiruchirappalli, the Department of Biotechnology and Microbiology, National College, Tiruchirappalli, and Saveetha Institute of Medical and Technical Sciences (SIMATS), Chennai, for providing the laboratory facilities and infrastructure necessary to carry out this research. The authors further appreciate the valuable contributions and dedication of all members of the research team involved in this study.
PGJ: Conceptualization, Methodology, Investigation, Formal analysis, Data curation, Writing—original draft, Writing—review & editing, Supervision. GA: Investigation, Formal analysis, Writing—review & editing. PAP: Methodology, Investigation, Data curation. RP: Formal analysis, Validation, Data curation. MH: Formal analysis, Validation, Data curation. MMJ: Conceptualization, Methodology, Resources, Funding acquisition, Supervision, Project administration, Writing—review & editing. All authors read and approved the submitted version of the manuscript.
The authors declare that they have no conflicts of interest.
The animal study was approved by the Institutional Animal Ethics Committee of National College (Autonomous), Tiruchirappalli, India (CPCSEA Approval No. 1941/GO/RE/S/17/CPCSEA).
Not applicable.
Not applicable.
The datasets generated and/or analyzed during the current study are partly included in this published article. Additional data supporting the findings of this study are available from the corresponding author upon reasonable request.
This work was supported by the University Grants Commission (UGC), New Delhi, Government of India [F.No. 43-557/2014(SR)]. The funding agency had no role in the design of the study, collection, analysis, or interpretation of data; writing of the manuscript; or in the decision to submit the article for publication.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 65
Download: 4
Times Cited: 0
