 Review
Review

Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
Affiliation:
Department of Translational Research, College of Osteopathic Medicine of the Pacific, Western University of Health Sciences, Pomona, CA 91766, USA
Email: vrai@westernu.edu
ORCID: https://orcid.org/0000-0001-6286-2341
Explor Dig Dis. 2026;5:1005128 DOI: https://doi.org/10.37349/edd.2026.1005128
Received: March 09, 2026 Accepted: May 14, 2026 Published: July 09, 2026
Academic Editor: Tzi-Bun Ng, The Chinese University of Hong Kong, China
The article belongs to the special issue Helicobacter Pylori and Infection: Genomics, Diagnosis, Pathogenesis, Antibiotic Resistance, Microbiota, Cancer, Prevention and Therapeutics
Gastric mucosa-associated lymphoid tissue (MALT) lymphoma represents a distinctive low-grade non-Hodgkin B-cell malignancy and one of the clearest examples of infection-associated carcinogenesis. Accounting for 7–9% of all B-cell lymphomas, this disease demonstrates a unique therapeutic paradigm, as early-stage gastric MALT lymphoma achieves complete remission in approximately 60–90% of patients following Helicobacter pylori eradication alone. This response reflects the tumor’s dependence on chronic antigenic stimulation, though disease progression and treatment resistance are driven by genetic alterations, such as the t(11;18) translocation, that confer antigen-independent growth. This review synthesizes current knowledge on clinical presentation, diagnostic evaluation, immunopathogenesis, prognostic factors, and evidence-based management of gastric MALT lymphoma. This review discusses the molecular mechanisms underlying the transition from antigen-dependent to antigen-independent disease, emerging therapeutic strategies, including targeted molecular therapies and BTK inhibitors, and addresses ongoing challenges in diagnosis, treatment resistance, and surveillance. Additionally, the broader implications of gastric MALT lymphoma to better understand how chronic infection drives lymphomagenesis and how pathogen eradication can lead to regression of antigen-dependent lymphoma, with the aim of providing insights for other microbe-associated malignancies, have been discussed.
Mucosa-associated lymphoid tissue (MALT) lymphoma is a low-grade extranodal marginal zone B-cell lymphoma that accounts for 7–9% of all B-cell lymphomas and 40–50% of primary gastric lymphomas [1]. MALT lymphoma can present in the salivary glands, lung, or thyroid, but the most typical location is the stomach [2]. Gastric MALT lymphoma pathogenesis has a strong association with Helicobacter pylori (H. pylori), and eradication of the pathogen is associated with complete histological response of the lymphoma in 60–90% of patients [1]. This disease predominantly affects middle-aged adults, with a mean diagnostic age of 56.9 years [3]. Interestingly, extra-nodal MALT lymphoma has shown a slight female predominance (54% female), although gastric MALT lymphoma shows a more balanced or slightly male-leaning predominance [4]. Over the past two decades, the overall incidence rates of MALT lymphoma have declined by approximately 1.93% annually within the United States [5]. However, racial disparities exist as incidence rates have remained stable in Black populations, despite declines in White, Hispanic, and Asian/Pacific Islander populations, potentially reflecting disparities in access to diagnosis, treatment, or underlying infection burden [5].
MALT lymphoma was first described in 1984 by Isaacson and Wright [6] using immunohistochemical analysis on multiple lesions sharing a common pattern of histogenesis. MALT-derived lymphomas were described clinically to remain localized for prolonged periods of time and thus be responsive to localized therapies. Additionally, the tumor samples were characterized by a noninvasive lymphoplasmacytic infiltrate in which follicle center cells can be seen invading epithelial structures, forming lymphoepithelial lesions [6]. However, it wasn’t until 1998 that the critical association between gastric MALT lymphoma and H. pylori was established, when Zucca et al. [7] demonstrated that lymphomas arose from B-cell clones at sites of chronic H. pylori-associated gastritis. This discovery fundamentally transformed the understanding of infection-driven carcinogenesis, and gastric MALT lymphoma has become established as a model for antibiotic therapy as a potential cancer cure [8].
Gastric MALT lymphoma serves as a paradigm for infection-associated malignancy, with several unique features that distinguish it from other cancers. For example, this disease demonstrates an indolent clinical course with excellent long-term prognosis, with overall survival (OS) rates of 89.4% and 74.4% at 5 and 10 years, respectively [9]. Beyond its clinical importance, gastric MALT lymphoma offers insights that may inform approaches to other microbe-associated malignancies, such as ocular adnexal MALT lymphoma associated with Chlamydia psittaci (C. psittaci), cutaneous MALT lymphoma linked to Borrelia burgdorferi (B. burgdorferi), and possibly other infection-driven cancers [10]. Therefore, understanding the mechanisms by which chronic antigen stimulation promotes lymphomagenesis and how eradication of that stimulus can reverse malignant transformation has wider implications for cancer prevention and treatment approaches.
H. pylori, a gram-negative spiral-shaped bacterium, is the most common chronic bacterial infection in humans, with a global prevalence of more than 40% [11]. It is the leading cause of infection-associated cancer and has been categorized as a group I (definite) carcinogen by the World Health Organization International Agency for Research on Cancer due to its causal association with gastric cancer [12]. While the global prevalence of H. pylori in adults has declined from 52.6% (before 1990) to 43.9% (2015–2022), similar trends have not been seen in children and adolescents [13]. Additionally, H. pylori disproportionately affects racial and ethnic minority groups. In a large cohort study of 913,328 individuals in the United States, H. pylori was diagnosed in 25.8% overall, with the highest positivity amongst non-Hispanic Black (40.2%) and Hispanic (36.7%) individuals, while non-Hispanic White individuals had the lowest positivity (20.1%) [14]. This highlights that although overall rates of H. pylori have decreased in recent decades, persistent disparities remain.
Infection is typically acquired in early childhood through intrafamilial horizontal transmission via gastric-oral or fecal-oral routes, establishing chronic gastric colonization in those who fail to clear the infection [12]. The bacterium is able to colonize the harsh acidic gastric environment through unique adaptations, such as its spiral morphology, chemotactic orientation, and formation of clonal population islands within gastric glands [15]. Urease is the most critical virulence determinant for H. pylori and is essential for both initial colonization and maintenance of chronic infection [15]. Urease enables the bacterium’s survival by hydrolyzing urea to ammonia and carbonic acid, neutralizing the gastric acid in the bacterium’s immediate microenvironment [16]. Additionally, urease has immunomodulatory roles and has been shown to induce regulatory T-cell (Treg) responses that promote immune tolerance and prolong neutrophil lifespan and subsequent chronic inflammation [17, 18].
The cytotoxin-associated gene pathogenicity island (cag PAI) also represents an imperative component of disease severity. This genetic element encodes a type IV secretion system (T4SSS) that forms a syringe-like pilus used to inject the cytotoxin-associated gene A (CagA) oncoprotein and other effectors into the host gastric epithelial cells [19]. Importantly, CagA plays a role in nuclear factor of activated T-cells cytoplasmic 1 (NFATc1) activation, which subsequently induces G1 cell cycle arrest and results in the characteristic slow-growing phenotype of MALT lymphoma. Nuclear NFATc1 localization is significantly associated with CagA presence and correlated with antibiotic responsiveness [20]. Patients exhibiting both CagA and nuclear NFATc1 have also demonstrated more rapid complete remission (CR) following H. pylori eradication (HPE) (median 4.00 vs. 6.00 months; p = 0.003) [20].
Beyond CagA, other H. pylori virulence factors likely contribute to the inflammatory and immune-modulating environment that promotes MALT lymphogenesis. For example, the vacuolating cytotoxin A (VacA) protein is a toxin that enters host cells and forms transmembrane pores in both the plasma membrane and the organelle membranes, resulting in disturbances to the homeostasis of ion exchanges within the cell. These alterations are then thought to disrupt epithelial lysosomes, endosomes, and mitochondria, facilitating successful invasion of the gastric epithelium [21]. Outer membrane proteins (OMPs), including lipoproteins, porins, and adhesions, also contribute to H. pylori adherence, invasion, and immune evasion. Specifically, the OMPs, OipA, SabA, and BabA enhance gastric mucosa colonization. Additionally, the OMP, HopH gene is associated with gastric mucosa inflammation [22, 23]. All strains of H. pylori also include neutrophil-activating protein (NAP), which enhances penetration and generation of oxygen radicals, further contributing to long-term inflammation within the gastrointestinal epithelium [23]. Interestingly, the cytokine expression profile differs between H. pylori-infected and non-infected gastritis. In H. pylori-positive (HPP) gastritis, interleukin (IL)-1β, IL-6, and IL-8 are paradoxically lower compared to H. pylori-negative (HPN) gastritis, while IL-12, IL-18, and tumor necrosis factor alpha (TNF-α) are elevated [24]. This distinct cytokine profile represents a unique inflammatory response involved in Th1 polarization and Treg activation [24, 25]. Chronic H. pylori infection also induces the formation of organized lymphoid follicles within the gastric mucosa [26]. This acquired MALT provides the substrate for subsequent lymphomagenesis in susceptible individuals. The inflammatory microenvironment is characterized by a Treg-balanced environment that appears to be a critical step in gastric MALT lymphoma development [27].
This environment is also characterized by a proliferation-inducing ligand (APRIL)-producing eosinophil, which targets tumoral B-cells, encouraging a pro-tumorigenic fate [27]. Additionally, the interplay between IL-17, IL-10, and APRIL production contributes to a complex cytokine network that sustains chronic inflammation and promotes B-cell proliferation and survival [24, 27]. H. pylori also employs immune evasion strategies, such as altering pattern recognition receptor signaling, inducing dendritic cell tolerance, and impairing phagocytic killing via interruption of the reactive oxygen species (ROS) and nitric oxide production [28]. Together, these inflammatory and immune-modulating effects create a microenvironment that promotes B-cell activation and survival, setting the stage for the molecular events that drive gastric MALT lymphomagenesis.
Persistent colonization of H. pylori results in chronic active inflammation of the gastric mucosa, recruitment of immune cells, and B and T lymphocytes, and the development of lymphoid follicles, creating ectopic MALT in sites typically devoid of organized lymphoid tissue [29] (Figure 1). This is due to increased secretion of prostaglandins (PGs, mainly PGE2) contributing to chronic inflammation by increased recruitment of immune cells secreting pro-inflammatory cytokines (Figure 1). Increased immune response causes death of parietal cells, leading to chronic atopic gastritis and intestinal metaplasia (Figure 1). The resulting environment sustains a continuously proliferating population of B-cells under ongoing antigenic and cytokine stimulation. DNA damage ensues secondary to neutrophil-derived ROS and H. pylori-induced endonucleases, promoting accumulation of genetic mutations in this constitutively stimulated B-cell compartment [30]. However, the relative contribution of ROS-mediated damage versus enzymatic DNA cleavage remains incompletely resolved, and these mechanisms may act synergistically within spatially distinct tumor niches, contributing to intratumoral molecular heterogeneity. Further genetic abnormalities, such as inactivation of the tumor suppressor gene p53, can lead to high-grade transformation to DLBCL [31].
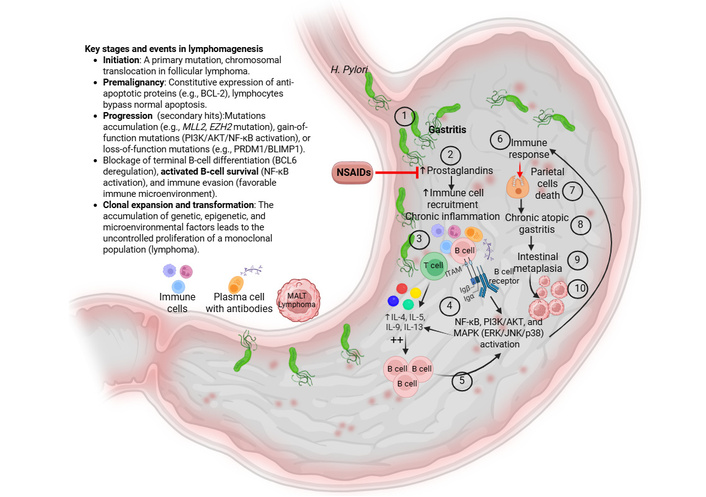
Etiology and key events in the pathogenesis of MALT lymphoma. Gastritis induces chronic inflammation when persistent irritants—primarily H. pylori infection, long-term NSAID use, or autoimmune response continuously damage the stomach’s protective mucosal barrier. This enduring injury (#1 and 2 in the figure) prevents healing, causing immune cells (lymphocytes/plasma cells) to infiltrate the stomach lining (#3 in the figure), leading to long-term mucosal damage and potential atrophy and replacement of gastric tissue with intestinal-type cells (intestinal metaplasia) (#4–10 in the figure). Increased recruitment of immune cells contributes to activation of inflammatory pathways, including NF-κB, PI3K-AKT signaling, and MAPK signaling, leading to increased secretion of pro-inflammatory cytokines. This leads to increased recruitment of B-cells with increased survival due to ongoing mutations. Clonal expansion and transformation of these cells result in an increased monoclonal population of these cells forming lymphoma. Red color indicates inhibition or negative effects. H. pylori: Helicobacter pylori; IL: interleukin; MALT: mucosa-associated lymphoid tissue; MAPK: mitogen-activated protein kinase; NF-κB: nuclear factor kappa B; PI3K: phosphatidylinositol 3'-kinase. Created in BioRender. Rai, V. (2026) https://BioRender.com/ouu5lje.
Tumor B-cell growth depends on H. pylori-specific intratumoral CD4+ T-cells, which provide help via CD40/CD40L co-stimulation and soluble ligands [29, 32]. In vitro studies demonstrate lymphoma B-cell proliferation in the presence of H. pylori-specific T-cells, with impaired growth when CD40/CD40L interactions are disrupted [33]. Most intratumoral T-cells exhibit a T-helper type 2 (Th2) phenotype, producing IL-4, IL-5, IL-13, and related cytokines inducing B-cell proliferation, survival, and class switching [29, 33, 34] (Figure 1). H. pylori infection also induces B-cell lymphoma production of chemokines including C–C motif chemokine ligand 17 and 22 (CCL17 and CCL22), recruiting Th2 cells and Tregs and fostering a tumor-supportive microenvironment [35, 36]. Conflicting reports of Th1-skewed responses suggest that dynamic Th1-Th2 plasticity, rather than a fixed phenotype, may underlie variability in disease behavior.
Gastric MALT lymphoma pathogenesis is also involved in infiltration of the lamina propria by CD4+, CD25+, FOXP3+ Tregs recruited by tumor-derived CCL17 and CCL22, suppressing host immunity and promoting persistent H. pylori infection [35, 37]. Murine models show that Treg eradication inhibits tumor growth comparably to total CD4+ T-cell depletion [35]. However, this remains controversial, as Tregs may also limit excessive inflammation; their net effect may depend on disease stage and cytokine context. Concurrently, APRIL-producing inflammatory cells, including eosinophils and tumor-associated macrophages (TAMs), provide potent B-cell survival signals and are enriched in H. pylori-driven lesions [27, 32]. Together, Th2 cells, Tregs, and APRIL-mediated signaling sustain antigen-dependent, apoptosis-resistant B-cells.
Beyond T-cell mechanisms, H. pylori strains expressing CagA can directly transform B-cells. CagA is translocated into gastric B lymphocytes via T4SS, undergoes tyrosine phosphorylation, and binds Src Homology 2 (SH2)-domain proteins (notably SHP-2), activating ERK and p38 mitogen-activated protein kinase (MAPK) pathways to promote aberrant B-cell proliferation and survival [29, 36, 38]. CagA also upregulates anti-apoptotic protein Bcl-2 and Bcl-xL and modulates p53 and Bcl-2-associated death promoter (BAD) phosphorylation, suppressing apoptosis [29, 36]. Variability in CagA expression and signaling suggests that host-dependent factors, including epigenetic susceptibility, may modulate its oncogenic effects. These mechanisms correlate with H. pylori-dependent MALT lymphomas and early DLBCL that regress after eradication therapy.
Tumor B-cell receptors (BCRs) in gastric MALT lymphoma are somatically mutated and polyreactive, recognizing diverse self and foreign antigens, including H. pylori antigens, IgG, DNA, and gastric components [39]. These BCRs receive both tonic and antigen-driven signals, and lymphoma B-cells proliferate upon stimulation with cognate antigens [35, 39]. This enables local autoantigens and microbial antigens to cooperatively sustain B-cell activation and survival. Such polyreactivity may also promote clonal diversification, allowing the emergence of subclones with differential antigen dependence during disease progression.
As disease progresses, accumulation of genetic lesions renders tumor growth antigen-independent and resistant to HPE. Recurrent translocations, including t(11;18)(q21;q21)/API2-MALT1, t(1;14)(p22;q32)/BCL10-immunoglobulin heavy chain (IGH), t(14;18)(q32;q21)/IGH-MALT1, and t(3;14)/FOXP1-IGH, converge on constitutive nuclear factor kappa B (NF-κB) activation via the CBM (CARMA/BCL10/MALT1) complex, driving proliferation and anti-apoptotic signaling independent of upstream BCR/T-cell input [38]. Lymphomas harboring t(11;18) or advanced NF-κB lesions typically fail to regress after HPE and may disseminate or transform to high-grade DLBCL [29, 30, 40]. Epigenetic alterations, including aberrant DNA methylation, may cooperate with these translations to stabilize NF-κB signaling and promote clonal evolution, though their temporal sequence remains unclear. This is particularly relevant in HPN MALT lymphomas, where antigen-independent growth may reflect earlier acquisition of such genetic and epigenetic lesions, suggesting a distinct yet convergent pathogenic pathway.
Similarly, MALT lymphomas driven by other infectious agents, such as C. psittaci and B. burgdorferi, share a common paradigm of chronic antigenic stimulation and T-cell-dependent B-cell activation, although they arise in distinct anatomic sites (e.g., ocular adnexa and skin) and may exhibit differing reliance on specific microbial-host signaling interactions [40, 41]. Notably, compared to H. pylori-associated gastric MALT lymphoma, these entities may demonstrate variable responsiveness to antimicrobial therapy, further underscoring both shared and divergent mechanisms of lymphomagenesis across infection-associated MALT lymphomas.
Gastric MALT lymphoma is typically an indolent, H. pylori-driven primary gastric lymphoma that often presents with nonspecific upper gastrointestinal symptoms and subtle endoscopic abnormalities, making a structured diagnostic workup essential for accurate diagnosis. The majority of patients initially present in mid-to-late adulthood with epigastric pain, dyspepsia, bloating, or recurrent nausea, often indistinguishable from benign H. pylori gastritis [42–44] (Figure 2). Less commonly, patients may present with weight loss, anemia, fatigue, and other symptoms [42]. Overt gastrointestinal bleeding in the form of melena or hematemesis is a rare complication but has been documented, including massive bleeding as the presenting event in HPN disease [44]. In pediatric patients, gastric MALT lymphoma presents similarly with abdominal pain, nausea, weight loss, and iron-deficiency anemia as some of the most common manifestations [45] (Figure 2).
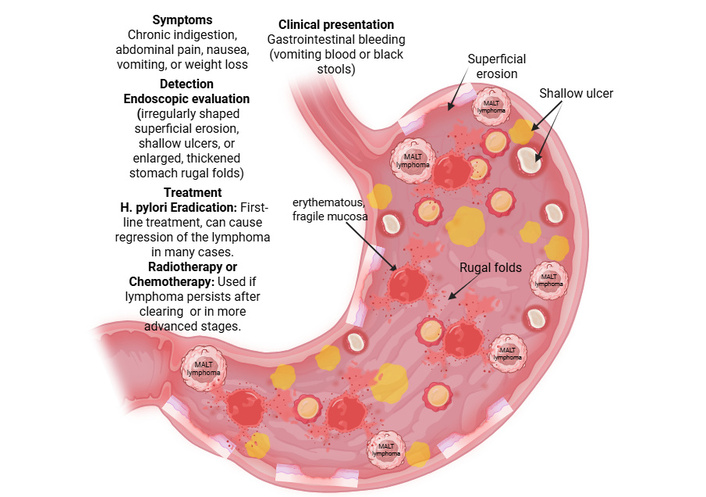
Symptoms, clinical presentation, diagnosis, and treatment for gastric MALT lymphoma. It presents with non-specific symptoms, most commonly chronic indigestion, abdominal pain, nausea, vomiting, or weight loss. The disease is often localized, with an excellent prognosis, though it can cause gastrointestinal bleeding. Endoscopy reveals erythematous, fragile mucosa, shallow ulcers, superficial erosion, and rugal folds in the stomach. Treatment primarily focuses on eradicating H. pylori with antibiotics in HPP cases. For cases unresponsive to antibiotics or HPN, localized radiotherapy or chemotherapy is used, offering high survival rates. H. pylori: Helicobacter pylori; HPP: H. pylori-positive; HPN: H. pylori-negative; MALT: mucosa-associated lymphoid tissue. Created in BioRender. Rai, V. (2026) https://BioRender.com/7hurhma.
Endoscopic identification of H. pylori-driven gastric MALT lymphoma can be complicated by its heterogeneous and often nonspecific appearance. Most frequently, endoscopy will reveal superficial erosions, small ulcers, or erythematous, fragile mucosa [42, 44, 45] (Figure 2). Less common endoscopic manifestations include thickened mucosal folds, nodularity, flat depressed/discolored lesions, and subepithelial tumor-like masses [44]. Alternatively, HPN and non-H. pylori Helicobacter (NHPH)-associated cases more often present on endoscopy as mass-like, proximal, non-superficial, or nodular gastritis-like lesions when compared with classic HPP cases [46]. Due to the high variability in appearance on endoscopy, histologic confirmation via biopsy is mandatory to confirm the diagnosis in any suspicious lesion.
Formal diagnosis of gastric MALT lymphoma requires multiple gastric mucosal biopsies from abnormal and mapping sites. Fine needle aspiration (FNA) alone is inadequate for diagnosis because tissue architecture (expansion of the marginal zone and presence of germinal centers) and lymphoepithelial lesions must be evaluated to rule out benign reactive processes such as chronic inflammation resulting from H. pylori gastritis [43, 47]. Histology shows the presence of marginal zone B-cells (monocytoid cells), lymphoepithelial lesions, and often background infiltration of chronic active H. pylori gastritis [42, 47]. Gastric MALT lymphoma typically presents with an immunophenotype of CD20+, CD79a+, BCL2+, CD5–/CD10–, with light-chain restriction; clonality can be supported by IGH rearrangement on molecular studies [44, 47–49]. Flow cytometry, when material allows, further documents a monotypic B-cell population [50]. Additionally, current guidelines recommend that HPP patients undergo FISH testing for t(11;18), which can confer antigen-independent growth; however, this is not universally performed [51, 52].
Because both prognosis and treatment hinge on infection status, rigorous H. pylori assessment is recommended utilizing two or more of the following methods: histology with special stains (Giemsa or Warthin-Starry), rapid urease test, urea breath test, stool antigen, or serology [42, 46, 49]. Comprehensive testing is considered standard in modern series given the high prevalence (~70–80%) of H. pylori positivity in gastric MALT lymphoma, although HPN cases now represent ~20–25% and are increasing [42, 46, 49]. Guidelines for defining MALT lymphoma as “HPN” require concordant negative results on at least two different H. pylori tests [46, 49]. HPN MALT lymphomas are more common in male patients, localize more proximally in the stomach, are mass-forming, and present with greater submucosal invasion compared to HPP disease [46]. NHPH-induced gastric MALT lymphoma linked to other bacterial species such as Helicobacter heilmannii (H. heilmannii) tends to present as a nodular gastritis-like pattern showing larger, more protruded, heterogeneous nodules in the angulus/body of the stomach, setting it apart from the typical H. pylori-induced nodular gastritis localized to the antrum [53].
Current staging systems for gastric MALT lymphoma aim to discern whether the disease is limited to the stomach and regional lymph nodes or has disseminated. The Lugano staging system is a modified version of the Ann Arbor system, which serves as the standard in contemporary series and radiotherapy cohorts for primary gastrointestinal lymphomas [42, 54, 55]. Staging is designated as follows: Stage I disease, which is confined to the gastrointestinal tract, Stage II disease, which extends into the abdomen, Stage II1 marked by local nodal involvement, Stage II2 marked by distant nodal involvement, Stage III, in which disease is localized to the gastrointestinal tract but with direct extension into adjacent organs, and Stage IV, which signals disseminated disease [56]. Workup to determine staging typically includes a computed tomography (CT) scan or positron emission tomography (PET) scan of the chest-abdomen-pelvis to assess nodes and extranodal spread, endoscopic ultrasound (EUS) to assess depth of wall invasion and regional nodes, and occasionally a bone marrow biopsy in rare instances where marrow involvement is suspected [43, 54].
Gastric MALT lymphoma typically has an excellent prognosis, but outcomes vary based on clinical, molecular, and H. pylori-related factors. The MALT Lymphoma International Prognostic Index (MALT-IPI) is a prognostic tool used to identify MALT lymphoma patients at risk of poor outcomes. Prognosis is based on three parameters: age greater than 70 years, Ann Arbor Stage III–IV, and elevated lactate dehydrogenase (LDH) level [57]. The MALT-IPI stratifies patients into low-, intermediate-, and high-risk groups with 5-year event-free survival (EFS) of about 70%, 56%, and 29%, respectively [57]. In contemporary gastric cohorts, most patients are classified within the low-risk group, reflecting localized, indolent disease [58, 59].
The dominant molecular determinant of gastric MALT lymphoma progression is the t(11;18)(q21;q21)/BIRC3-MALT1 translocation, which results in activation of the NF-κB pathway, promoting cell survival [60]. The translocation occurs in ~20–25% of gastric MALT lymphomas and is enriched in both HPN and disseminated cases [60]. It is also strongly associated with H. pylori-independent growth and resistance to HPE therapy, with one large series finding t(11;18) present in 42 of 63 non-responders, but only 2 of 48 complete responders [60]. Patients with the translocation experienced shorter progression-free survival (PFS) or EFS, although OS is often preserved with appropriate second-line therapy [51, 60]. Extra copies of the MALT1 gene, as seen in trisomy disorders or tetrasomy 18, also predict poorer EFS and greater risk of tumor progression or relapse, especially in advanced-stage (Stage IV) gastric MALT lymphoma. Specifically, this genetic aberration is associated with increased resistance to HPE, higher rates of progression or relapse, and lower EFS [61, 62]. Other recurrent alterations such as t(1;14) translocation, trisomy 3/7/12, and loss of TNF-α-induced protein 3 (TNFAIP3/A20) result in increased activation of the NF-κB pathway; at present, these have limited routine prognostic use but may influence response to targeted agents [60].
Multiple clinical and molecular series show that the presence of t(11;18) translocations in tumor cells confers resistance to tumor regression upon HPE. Translocation-positive tumors rarely achieve CR with antibiotic use alone, with most requiring treatment with radiotherapy or systemic therapy [58]. A higher incidence of t(11;18) translocation has been found among HPN gastric advanced -stage MALT [63].
The strongest clinical prognostic factors of gastric MALT lymphoma are depth of tumor invasion, regional lymph node involvement, H. pylori status, and tumor location. Depth of tumor invasion is determined via EUS, and the presence of submucosal (or deeper) invasion strongly predicts poor response to HPE, higher likelihood of need for radiotherapy or systemic therapy, and more aggressive disease behavior [54]. Involvement of regional lymph nodes (Lanugo Stage II) is associated with lower locoregional relapse-free survival (LRFS) after radiotherapy (Stage IIE vs. IE) and decreased response to eradication as well as a more “aggressive-course” classification in modern cohorts [58]. H. pylori infection status carries both a predictive and prognostic impact on the disease course. Patients who are HPP with localized disease have excellent CR rates (~70–80%) and long-term outcomes with eradication alone [42]. Alternatively, HPN status is consistently an adverse factor associated with worse PFS as seen in a large real-world cohort, as well as higher rates of advanced stage, proximal or multifocal lesions and the need for radiotherapy/chemotherapy [42]. Tumor location is predictive of tumor response to HPE as well as disease prognosis. Lesions localized to the proximal or upper-third portion of the stomach and tumors in multiple gastric locations are independently associated with poor response to HPE and higher cumulative incidence of relapse and progression [42]. Conversely, as seen in most large series of typical HPP MALT lymphoma, disease localized to the distal/antral superficial mucosa of the stomach typically has excellent outcomes [58].
Characterization of prognosis in H. pylori-associated gastric MALT lymphoma is best characterized by combining various prognostic indicators (global index, clinical factors, and genetics) into an integrated risk assessment. At present, prognosis is determined using the MALT-IPI (age, stage, LDH), tumor localization within the stomach, and depth of invasion, nodal involvement, H. pylori positivity, and presence or absence of t(11;18) translocation and extra MALT1 copies [57]. Patients with MALT-IPI categorization of low risk, HPP, distal, mucosal-limited, t(11;18)-negative disease have excellent long-term survival and high cure rates with HPE alone, whereas HPN, t(11;18)-positive, proximally located, deeply invasive, or node-positive tumors are more likely to be refractory to remission via antibiotic eradication and require radiotherapy or systemic therapy, with shorter PFS despite generally preserved OS [58].
Gastric MALT lymphoma represents a unique paradigm in oncology, as one of the only malignancies that can be cured by treating the underlying infection rather than the cancer itself. Following successful eradication, CR occurs in approximately 75.18% of patients (95% CI: 70.45–79.91%) with early disease [55] (Figure 3). Treatment typically consists of a proton-pump inhibitor alongside amoxicillin and either clarithromycin or metronidazole [1], though strategies must be tailored to H. pylori status, disease stage, gene aberrations, and response to initial therapy [64] (Figure 2).
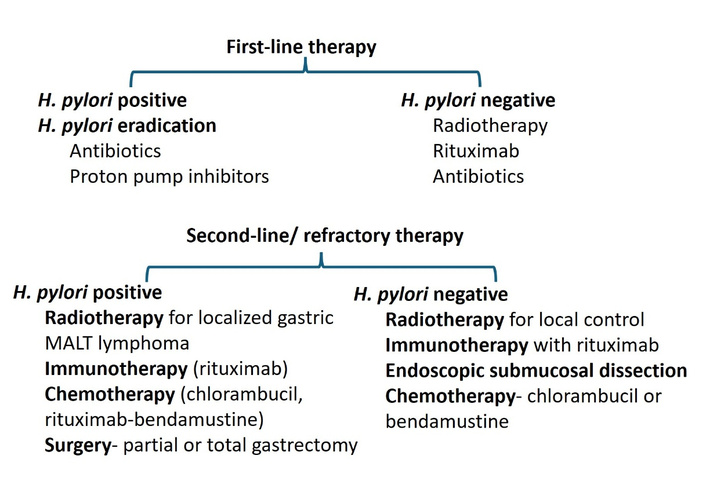
First- and second-line therapy for HPP and HPN gastric MALT lymphoma. H. pylori: Helicobacter pylori; HPP: H. pylori-positive; HPN: H. pylori-negative; MALT: mucosa-associated lymphoid tissue.
For HPN gastric MALT lymphoma, the role of antibiotics remains debated. A 2024 retrospective analysis found CR rates of 63% in HPP patients versus 13% in HPN patients (p = 0.033), suggesting HPE is not an effective first-line treatment for HPN disease [65]. In contrast, Gong et al. [66] concluded that eradication therapy is worthwhile regardless of H. pylori status, while a 2021 retrospective study of 178 patients favored radiation therapy (RT) as the standard of care due to its favorable outcomes, with 5- and 10-year OS rates of 94% and 79%, respectively [67] (Table 1). Notably, some HPN cases may harbor other organisms such as H. heilmanni or H. felis, which may respond to clarithromycin-based regimens [68].
Summary of the results of previous studies for gastric MALT lymphoma.
| Study | Country | Cases | H. pylori status | Genetic alternations | Clinical associations | Strengths/Limitations |
|---|---|---|---|---|---|---|
| Gong et al., [66] | South Korea | 345 | HPP + HPN | NR | Suggested HPE may be attempted initially regardless of H. pylori status | Retrospective review; variable diagnostic methods |
| Laoruangroj et al., [65] | Multicenter | 52 | HPP vs. HPN | NR | CR: 63% in HPP vs. 13% in HPN, showing reduced efficacy of HPE in true HPN disease | Direct comparison; retrospective |
| Olszewski et al., [69] | USA (SEER-Medicare) | 1,134 | Mixed | NR | Comparative outcomes across therapies support the effectiveness of RT and systemic therapy in appropriate patients | Population-based, retrospective, limited molecular data |
| Yahalom et al., [67] | USA | 178 | HPN | NR | RT showed excellent outcomes in HPN disease; 5- and 10-year OS ~94% and 79% | Large HPN cohort; retrospective |
| Jeong et al., [70] | South Korea | 229 | HPE-resistant/HPN | NR | RT achieved durable control; 10-year OS ~92.8% with low toxicity | Long follow-up; retrospective |
| Jeon et al., [71] | South Korea | 62 | HPE-resistant/HPN | NR | Reduced-dose RT (24–25.5 Gy) achieved ~96.7% CR at 6 months and ~92% 5-year local control with minimal toxicity | Prospective design; strong evidence for RT dosing |
| Opat et al. (MAGNOLIA Trial), [72] | Multicenter | 68 | Mixed | NR | Zanubrutinib demonstrated ORR ~68.2% with durable responses in relapsed/refractory MZL | Prospective clinical trial; not gastric-specific |
CR: complete remission; H. pylori: Helicobacter pylori; HPE: H. pylori eradication; HPN: H. pylori-negative; HPP: H. pylori-positive; MALT: mucosa-associated lymphoid tissue; OS: overall survival; RT: radiation therapy.
Radiotherapy (24 Gy) directed at the stomach and perigastric nodes provides effective disease control in patients with localized HPN lymphomas or in patients resistant to HPE therapy [73] (Figures 2 and 3). A 2025 retrospective analysis found that the LRFS, disease-free survival (DFS), and OS rates at 10 years were 90.3%, 87.7%, and 92.8%, respectively, with low toxicity [70]. Similarly, a 2026 study of Stage I–II gastric MALT lymphoma with resistance to HPE therapy demonstrated that radiotherapy alone was associated with superior PFS (p = 0.001) and OS (p = 0.031) compared to systemic therapy, with no added benefit from combination therapy [74]. Prospective data confirms that reduced-dose RT (24–25.5 Gy) achieves high CR rates (96.7% at 6-months) with excellent local control and minimal toxicity, supporting dose de-escalation strategies [71] (Table 1). For patients in whom involved-site radiotherapy (ISRT) is contraindicated or unavailable, rituximab monotherapy (375 mg/m2 weekly for four doses) represents an effective alternative, achieving CR with minimal toxicity [75].
For patients with persistent lymphoma after HPE, the National Comprehensive Cancer Network (NCCN) recommends management dependent on symptom status and disease behavior. Asymptomatic patients with stable disease may undergo repeat endoscopy and biopsy after 3 months or proceed to ISRT, while symptomatic or progressive disease warrants ISRT [76] (Figure 3). A watch-and-wait strategy is safe for patients with minimal histological residuals, as 94% demonstrate favorable disease courses without oncological treatment [76].
When systemic therapy is required, rituximab plus chlorambucil achieved 92% CR and 100% overall response at 2-years, with a 5-year PFS of 89%, significantly better than rituximab alone (64% CR, 70% PFS) or alkylating agents alone (66% CR, 68% PFS) [77]. For relapsed or refractory disease, Bruton’s tyrosine kinase (BTK) inhibitors like zanubrutinib demonstrated a 68.2% ORR and an estimated 15-month PFS of 82.5%, leading to FDA approval [72]. Rituximab plus lenalidomide is also approved, and ibrutinib demonstrated a 58% ORR with 15.7-month PFS [78].
Active surveillance is recommended for asymptomatic patients with advanced-stage disease, as treatment delay does not impair survival [78]. Indications for treatment include gastrointestinal bleeding, threatened end-organ function, bulky disease, or rapid progression [78]. When treatment is required, ISRT is preferred if the disease can be encompassed within a single radiation field; otherwise, systemic therapy with rituximab alone or combined with chlorambucil or bendamustine is appropriate [78]. Patients with t(11;18)-positive disease require special consideration due to higher HPN virulence and resistance to antibiotic therapy [79]. Rituximab-chlorambucil was significantly more efficient than rituximab alone in these patients, achieving remission in 100% vs. 45% at week 6 (p = 0.0005) [80].
High-grade transformation to DLBCL occurs in approximately 1.59% of cases and requires immediate management with [81] anthracycline-based chemoimmunotherapy [73]. After multiple prior therapies, chimeric antigen receptor (CAR) T-cell therapy, bispecific antibodies, or clinical trials may be considered [73, 82]. Surgery is recommended for massive bleeding or perforation [83].
Extended follow-up with endoscopy and tissue sampling is recommended after successful HPE, as complete regression may be prolonged and recurrence can occur with or without H. pylori infection [84]. One large-scale international study supports endoscopy every 3–6 months for the first 2 years, extending to every 6–12 months thereafter [84]. Most guidelines recommend a 24-month observation period before considering alternative treatment [85].
Early diagnosis of gastric MALT lymphoma remains difficult due to the nonspecific nature of presenting symptoms [86]. Clinical manifestations are vague and varied, including abdominal pain, dyspepsia, and weight loss being common presenting complaints that overlap substantially with benign gastric conditions [84]. The endoscopic appearance of gastric MALT lymphomas also varies, ranging from subtle erosions to nodular masses, and can be infiltrative, exophytic, or ulcerative [86]. Furthermore, the tumor can have multifocal distribution, necessitating aggressive tissue sampling with multiple biopsies from both abnormal and normal-appearing mucosa for accurate diagnosis [86]. Additionally, FNA is inadequate for diagnosis since small lymphoma cells cannot be distinguished from reactive B-cells and tissue architecture cannot be examined [87]. However, the Wotherspoon score in gastric biopsies or the demonstration of monoclonality may help differentiate lymphoma from chronic inflammation, but this distinction remains challenging in the absence of adequate biopsy material [88]. Delayed diagnosis may result in disease progression, acquired treatment resistance, and potential transformation into aggressive DLBCL [53]. Therefore, developing novel endoscopic techniques for in vivo diagnosis and identifying serum biomarkers for early detection are critical steps in negating MALT lymphoma impacts [73, 85].
Another diagnostic challenge arises from the increasing incidence of HPN gastric MALT lymphoma, which now represents the majority of cases in some centers. The pathophysiology of HPN MALT lymphoma without chronic gastritis remains unclear, and optimal management strategies are still being researched [49]. This offers an important research subject that would benefit the increasing number of patients with HPN MALT lymphomas.
Another difficulty in treating gastric MALT lymphoma is the transition from an antigen-dependent to an antigen-independent cancer. In early-stage disease, the lymphoma remains dependent on chronic antigenic stimulation, both via direct autoantigens and via indirect H. pylori-specific intratumoral T-cells, resulting in clonal expansion and survival [29]. However, acquired dysregulation of molecular pathways can result in tumors that are no longer dependent on their microenvironment [73]. Transformed clones carrying the t(11;18)(q21;q21) translocation form a MALT lymphoma whose growth is independent of H. pylori and will not respond to bacterial eradication [89]. Additionally, later genetic events, such as t(1;14)(p22;q32) involving BCL10, appear to be linked to a capacity for autonomous growth, loss of sensitivity to H. pylori, and dissemination of the lymphoma beyond the stomach [90].
The t(11;18)(q21;q21) BIRC3/MALT1 translocation is strongly associated with resistance to HPE [91]. Approximately 25% of patients harbor the t(11;18)(q21;q21) translocation, which has been associated with significantly lower 10-year PFS (26%) when compared with those without the translocation (57%; p = 0.004) [51]. Other factors that predict impaired treatment response include deeper invasion beyond the mucosa and regional lymph node involvement, which are much less likely to respond to antibiotic therapy [42]. Involvement of regional lymph nodes and bone marrow infiltration at diagnosis is associated with a worse prognosis overall [78]. Additionally, HPN status predicts poor response to eradication therapy, with CR rates of only 13% compared to 63% in HPP patients [49]. Without upfront identification of treatment-resistant disease, patients may undergo ineffective antibiotic therapy for months before being escalated to radiotherapy or systemic treatment. Better understanding of the interaction among tumor genetics, the microenvironment, and the microbiota at different anatomical sites will help in the selection of targeted treatments and the upfront identification of biomarkers for early progression [73]. Similarly, circulating tumor DNA (ctDNA) shows a potential for risk stratification and molecular response modeling [92].
Concurrently, the optimal timing for escalation from watchful waiting to active therapy remains uncertain, particularly in asymptomatic patients with minimal histological residuals after HPE [1]. Follow-up studies have shown the persistence of monoclonal B-cells after histologic regression in many cases, as well as transient molecular or histologic relapses without clinical or endoscopic evidence of disease [73]. Additionally, the optimal surveillance interval has not been definitively established, and the risk of secondary malignancies, including gastric adenocarcinoma and non-Hodgkin lymphoma or high-grade transformation, necessitates indefinite endoscopic monitoring [85, 86]. Therefore, prospective studies establishing evidence-based surveillance intervals, identification of transformation-predictive biomarkers, and strategies for secondary malignancy prevention in high-risk populations are needed.
Recent work links H. pylori-driven signaling, NF-κB pathway activation, and the broader gastrointestinal microbiome to new therapeutic strategies for gastric MALT lymphoma. Constitutive NF-κB pathway activation has been identified as a central driver in HPN and t(11;18)-positive MALT lymphomas, via translocations and mutations in TNFAIP3 and related regulators [38, 93]. This has led to interest in direct NF-κB pathway inhibition, including next-generation proteasome inhibitors, especially in antibiotic-refractory disease [93, 94]. These findings have also inspired the development of MALT1 protease inhibitors, which sit at the convergence of BCR and NF-κB signaling, with preclinical agents showing potent activity and preliminary relevance to MALT1-rearranged gastric MALT lymphoma [93, 95]. Combining eradication therapy with targeted disruption of CBM-NF-κB signaling (CARD11-BCL10-MALT1) is a likely future direction for high-risk or eradication-refractory subsets [95–97].
Approaches to treatment are increasingly being refined through genetic profiling, microbiome-aware strategies, and predictive biomarkers. Next-generation sequencing (NGS) and targeted panels can characterize translocations (such as API2-MALT1) and NF-κB pathway mutations to identify patients unlikely to benefit from HPE alone for early RT or systemic therapy initiation [93]. BCR NGS (IgH/IgL) enables more sensitive detection and monitoring of clonal disease, facilitating earlier identification of minimal residual disease (MRD) and relapse [98]. Transcriptomic profiling of HPN, API2-MALT1-negative disease has yielded candidate genes including OLFM4 and NANOG as predictive markers of eradication response [93]. Broader frameworks such as liquid biopsy, ctDNA, and immune-microenvironment signatures are likely to be adapted to indolent lymphomas, including gastric MALT lymphoma, to personalize therapeutic intensity [99, 100].
H. pylori infection results in substantial gastric and intestinal dysbiosis, and other NHPH species and microbiome shifts appear relevant to gastric MALT lymphoma pathogenesis and treatment response [38, 101]. Gut microbiota composition modulates tumor response to chemotherapy, immunotherapy, and antibiotics and can be altered by diet, probiotics, prebiotics, and fecal microbiota transplantation [102–104]. Following HPE, only partial restoration of microbial diversity is seen, with persistent depletion of beneficial genera [102]. Future management may integrate microbiome profiling and interventions, including selective probiotics, high-fiber or anti-inflammatory diets, and judicious antibiotic use to maintain immune homeostasis and possibly reduce recurrence risk [101]. Experimental data showing that certain Lactobacillus strains can prevent Helicobacter-associated lymphoid lesions in animal models suggest a conceptual path toward probiotic-based prevention, though human data remains lacking [38].
Given H. pylori’s status as a class I carcinogen, future public health-oriented strategies may implement population-level screening and eradication in high-incidence regions, alongside addressing modifiable factors such as smoking, alcohol consumption, and high-salt diets that contribute to gastric pathology [38, 102, 103]. The rapid expansion of NGS, transcriptomics, and microbiome sequencing generates high-dimensional data ideally suited for artificial intelligence (AI)-driven analysis, which could integrate mutation profiles, gene-expression signatures, and microbiome composition to build predictive models of eradication response, transformation risk, or need for systemic therapy [93, 99, 104].
The success of HPE as first‑line, often curative therapy for early gastric MALT lymphoma (~70–80% CR in HPP localized disease) has become a paradigm for microbe‑targeted treatment of indolent lymphomas [55, 105]. This experience has directly inspired analogous strategies in other bacteria‑associated extranodal MZL/MALT lymphomas [41, 106]. In ocular adnexal MALT lymphoma, data linking C. psittaci to a subset of tumors led to trials of front‑line doxycycline, with a proportion of patients achieving durable lymphoma regression after antibiotic therapy alone [68, 106]. Similarly, Campylobacter jejuni-associated immunoproliferative small intestinal disease (IPSID) and some cases of intestinal MALT lymphoma respond to targeted antibiotic regimens [106]. For cutaneous MALT lymphomas, reports associating B. burgdorferi with disease and occasional responses after Borrelia‑directed antibiotics have further reinforced the concept of “infection‑driven”, reversible lymphoma [41, 106]. These observations support a broader future direction in which careful microbial work-up (including PCR/serology for organisms such as C. psittaci or Borrelia) could become standard in anatomically distinct MALT lymphomas, with trial antibiotic therapy considered as an initial, low‑toxicity option when a plausible infectious driver is identified [47, 68, 106]. More broadly, the gastric H. pylori model supports the concept that prevention or early treatment of specific chronic infections could reduce the incidence of several extranodal lymphomas [41, 106].
Despite advances in therapeutic approaches, gastric MALT lymphoma continues to present diagnostic and management challenges. The increasing proportion of HPN cases and the limited understanding of their biology complicate management decisions. Determining when persistent histologic or molecular disease after eradication warrants treatment escalation remains difficult. These uncertainties highlight the need for improved biomarkers, standardized surveillance strategies, and clearer thresholds for intervention. Transformation to diffuse large B-cell lymphoma, although uncommon, further underscores the importance of risk stratification and long-term follow-up.
APRIL: a proliferation-inducing ligand
BAD: Bcl-2-associated death promoter
BCRs: B-cell receptors
BTK: Bruton’s tyrosine kinase
CagA: cytotoxin-associated gene A
CAR: chimeric antigen receptor
CCL17: C–C motif chemokine ligand 17
CR: complete remission
ctDNA: circulating tumor DNA
DFS: disease-free survival
EFS: event-free survival
EUS: endoscopic ultrasound
FNA: fine needle aspiration
H. pylori: Helicobacter pylori
HPE: Helicobacter pylori eradication
HPN: Helicobacter pylori-negative
HPP: Helicobacter pylori-positive
IGH: immunoglobulin heavy chain
IL: interleukin
IPSID: immunoproliferative small intestinal disease
ISRT: involved-site radiotherapy
LDH: lactate dehydrogenase
LRFS: locoregional relapse-free survival
MALT: mucosa-associated lymphoid tissue
MALT-IPI: mucosa-associated lymphoid tissue Lymphoma International Prognostic Index
MAPK: mitogen-activated protein kinase
MRD: minimal residual disease
NCCN: National Comprehensive Cancer Network
NFATc1: nuclear factor of activated T-cells cytoplasmic 1
NGS: next-generation sequencing
NHPH: non-Helicobacter pylori Helicobacter
OMPs: outer membrane proteins
OS: overall survival
PET: positron emission tomography
PFS: progression-free survival
ROS: reactive oxygen species
RT: radiation therapy
TAMs: tumor-associated macrophages
Th2: T-helper type 2
TNFAIP3: tumor necrosis factor alpha-induced protein 3
TNF-α: tumor necrosis factor alpha
Treg: regulatory T-cell
SS: Conceptualization, Writing—original draft. CT: Conceptualization, Writing—original draft. VR: Writing—review & editing, Supervision. All authors read and approved the submitted version.
The authors declare that there are no conflicts of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 1882
Download: 31
Times Cited: 0
Luiza Gilmanovna Bektemirova ... Vasiliy Ivanovich Reshetnyak
Elias Kouroumalis ... Argyro Voumvouraki
Rosabel Corrales ... Rafael Llanes
Natalia V. Baryshnikova ... Alexander N. Suvorov
Tatyana Anatolievna Kuchmenko ... Arina Kopaeva
Surbhi Dumra, Abhishek Ray
Vasisht Karri ... Samir M. Dalia
