 Review
Review

Affiliation:
1Department of Chemistry, University of Wisconsin-Madison, Madison, WI 53706, USA
2Biosciences Center, National Laboratory of the Rockies, Golden, CO 80401, USA
†These authors contributed equally to this work.
ORCID: https://orcid.org/0009-0005-4942-0369
Affiliation:
2Biosciences Center, National Laboratory of the Rockies, Golden, CO 80401, USA
†These authors contributed equally to this work.
ORCID: https://orcid.org/0000-0002-3550-188X
Affiliation:
2Biosciences Center, National Laboratory of the Rockies, Golden, CO 80401, USA
Email: Yannick.Bomble@nlr.gov
ORCID: https://orcid.org/0000-0001-7624-8000
Explor BioMat-X. 2026;3:101367 DOI: https://doi.org/10.37349/ebmx.2026.101367
Received: February 18, 2026 Accepted: May 21, 2026 Published: June 02, 2026
Academic Editor: Håvard Jostein Haugen, University of Oslo, Norway
The article belongs to the special issue Nature-Based Biomaterials for Biomedical Applications
The development of biomaterials capable of supporting complex tissue growth remains a central challenge in regenerative medicine and tissue engineering, particularly in replicating the structural, mechanical, and transport functions of native extracellular matrices. While decellularized animal tissues have demonstrated significant success as scaffolds for tissue engineering, they are still constrained by cost, immunogenicity, and ethical concerns. In recent years, decellularized plant tissues have emerged as a compelling alternative scaffold platform due to their inherent vascular architectures, ethical sourcing, tunable mechanical properties, cytocompatibility, and sustainability. This review summarizes current strategies for the decellularization of plant tissues, including chemical, enzymatic, and physical approaches, and discusses how these methods preserve plant cell wall structure while removing immunogenic components. Advances in surface loading and functionalization, including protein coatings, oxidation, nanoparticle incorporation, peptide conjugation, and bioactive molecule loading, have further enhanced cell adhesion, differentiation, biodegradability, and immunomodulation. Recent applications of decellularized plant scaffolds in cardiac, skeletal muscle, bone, nerve, and wound healing contexts are reviewed, highlighting proof-of-concept successes and remaining challenges. Beyond therapeutic applications, plant-derived scaffolds have also enabled physiologically relevant in vitro models for vascular biology, mechanotransduction, cancer, metabolic tissues, and drug response studies. Collectively, these advances position decellularized plant tissues as versatile, low-cost, and ethically favorable biomaterials with growing relevance for both regenerative medicine and tissue modeling.
The creation of medically relevant biomaterials and artificial organs capable of replacing natural organs taken from transplant donors has long been a goal in the fields of regenerative medicine and tissue engineering. Stem cells possess unique self-renewal and differentiation capacities, and thus offer a potential route to organ engineering and regeneration [1]; however, maintaining healthy cell growth in tissues with a thickness greater than 100–200 µm (above the diffusion limit of oxygen), and achieving precise control over cellular fate specification and higher-order tissue organization remains a significant challenge [2]. Cells rely on a complex interplay of biochemical, temporal, positional, and biomechanical signals to correctly differentiate into the necessary cell type which are not easily recreated in vitro [3–5]. For example, mesenchymal stem cell differentiation is potently impacted by biomechanical signaling. When grown on substrates with an elastic modulus mimicking that of brain (~1 kPa), muscle (~10 kPa), or collagenous bone (~100 kPa), mesenchymal stem cell differentiation is directed towards the corresponding neurogenic, myogenic, or osteogenic lineage [6]. In vivo, these mechanical cues, essential structural support, and transport pathways for nutrients, waste, and signaling molecules are provided by a highly organized extracellular matrix (ECM), which is comprised of macromolecules such as elastin, collagen, fibronectin, laminin, and other matrix proteins [7]. Scaffolds for cellular growth therefore serve an essential role in recreating these important functions of the ECM to promote directional growth [8], improve differentiation [9, 10], facilitate the transport of nutrients and waste, and promote the vascularization of new tissue [2]. Scaffolds can be characterized as either natural scaffolds created from decellularized biological tissues or natural ECM derivatives (collagen, gelatin, chitosan, etc.) [11, 12], synthetic scaffolds which are generated from synthetic polymers such as polyglycolic acid (PGA) and polylactic acid (PLA) [13], or hybrid scaffolds which use a combination of both natural and synthetic ECM components [12]. Some of the early intellectual heritage to decellularized biological tissue scaffolds can be traced to work from the late 1970s where studies into the biosynthesis of glomerular basement membrane matrixes, a specialized type of ECM which acts as a barrier between urine and blood in the glomeruli capillaries, used washes with sodium deoxycholate to purify basement membrane matrixes by removing contaminating cell membranes in a manner reminiscent of some decellularization techniques used today [14]. Work in the mid-1980s advanced these capabilities even further by culturing rat pneumocytes on decellularized human lung acellular alveolar matrix fragments prepared by sonication and incubation of the lung tissue in sodium deoxycholate, effectively demonstrating the culture of non-native cells on a decellularized biological matrix [15]. Decellularization of full tissues was not demonstrated until 1995 when porcine small intestinal submucosa matrices were decellularized and used as a biomaterial to correct an induced Achilles tendon defect in dogs [16]. Since then, whole hearts, livers, lungs, kidneys, as well as numerous tissues have been successfully decellularized and in some cases seeded with fresh cells to regain some native function [17–20].
Although animal scaffolds have been tremendously successful, they still suffer from key issues such as high production costs, batch-to-batch variability, potential disease transmission to patients, xenogeneic differences in ECM composition that can trigger immune responses, and ethical concerns surrounding their production [21, 22]. Decellularized plant cells arose over the last decade to address some of these core drawbacks, with the earliest accounts of decellularized plant tissues being used as scaffolds to support mammalian cell growth originating from papers published by Modulevsky et al. in 2014 and 2016 [23, 24]. In these studies, the authors demonstrated that decellularized apple hypanthium tissues could support 3D mammalian cell growth and were biocompatible after implantation into immunocompetent mice. Combined with prior studies which established that synthetically produced cellulose scaffolds were capable of supporting limited 3D mammalian cells growth [25], the authors reasoned that the air pocket and pore rich structural composition of apple hypanthium could yield an easily produced scaffold for 3D mammalian cell culture. In 2017, Gershlak et al. [26] advanced the field further by taking advantage of the intact vasculature already present in decellularized spinach leaves to culture cardiomyocytes capable of eliciting contractile responses. Since then, a growing number of publications on decellularized plant scaffolds have shown that they possess remarkable potential as scaffolds to support tissue engineering efforts owing to their inherent vasculature and favorable mechanical properties which can mimic those of mammalian tissues [27], cytocompatibility, and low immunogenicity [24, 28]. For example, similarities between the hierarchical structure of bone and bamboo have been drawn in reference to both materials’ role as a high stiffness and high porosity material comprised of repeating monomers that combine into larger fibrous structures [29]. This observation, in conjunction its high interconnected porosity (≥ 90%), pore size appropriate for cellular and vascular infiltration (≥ 100 µm), and mechanical properties (~25 GPa elastic modulus) in the range found for cortical bone (10–30 GPa) [30] has made bamboo an attractive scaffold for bone tissue engineering [31, 32]. In this manner, the varying mechanical properties of different plant tissues after decellularization can be matched to a corresponding organ type with an ECM that possesses similar properties [27]. Altogether, plant materials possess real potential advantages as scaffolds for tissue engineering considering they are readily available, cheap, ethically unambiguous, possess inherent vasculature, and are easy to prepare. Beyond this, decellularized plant scaffolds have also benefited from advancements in scaffold chemical functionalization, which opens new possibilities for tuning scaffold mechanical properties, modulating scaffold degradation rates, and enabling controlled release of medicines to achieve specific therapeutic outcomes. Given their tremendous potential as novel biomaterials, this review will cover essential concepts pertaining to the use of decellularized plant tissues as novel biomaterials, such as decellularization techniques and methods for scaffold functionalization, and outline the recent developments regarding their applications as scaffolds in regenerative medicine and tissue engineering.
Decellularization is a critical step in the preparation of plant-derived scaffolds used in tissue engineering, as it removes immunogenic native cellular material while preserving the three-dimensional architecture and mechanical integrity of the underlying plant cell wall [33]. Plant tissues have highly diversified structures and mechanical properties [27, 34, 35], and this breadth of material properties has led to the development of a wide array of protocols for decellularizing plant tissues using chemical, enzymatic, and physical techniques with various benefits and drawbacks (see Figure 1) [36]. Early plant decellularization approaches mirrored protocols developed for animal tissues, relying on perfusion techniques or simple incubation of plant tissues in a bath of hexanes, to remove waxy plant cuticles, followed by chemical detergents such as the non-ionic surfactant Triton X-100 and sodium dodecyl sulfate (SDS) for 1 to 5 days to disrupt lipid bilayers, denature proteins, and effectively lyse cells to facilitate their removal from the polysaccharide-rich plant cell wall [23, 26]. The success of a decellularization procedure is commonly assessed by measuring the leftover quantities of DNA in the scaffold, with a satisfactory decellularization resulting in less than 50 ng of DNA present per mg of plant tissue [37, 38]. These detergent-based approaches were instrumental in the first demonstrations that intact plant tissues, such as spinach or parsley leaves, could be rendered acellular while preserving their native vascular networks and gross morphology, establishing proof-of-concept for plant-based scaffolds in tissue engineering [23, 26, 28]. While the use of ionic detergents is by far the most widely used decellularization method, detergent-free methods based on leaf “skeletonization” techniques have also been developed which use heated bleach and sodium bicarbonate baths to achieve decellularization [39]. Nuclease treatments have also been used to decellularize tobacco plant cells, rice cells, and tobacco hairy root matrices [40]; however, the use of other enzymatic treatments that explicitly degrade non-cell wall components of plant tissues, such as proteases or phospholipases [41], to reduce reliance on harsh detergents and decellularization conditions remains relatively underexplored. One study seeking to develop standardized decellularization protocols on leatherleaf viburnum compared the use of SDS to a mixture of tergitol and trypsin, a digestive enzyme which cleaves peptides at the C-terminus of arginine and lysine residues, finding that SDS with a clearing solution (10% sodium hypochlorite, 0.1% Triton X-100) was overall more efficient at decellularization than tergitol and trypsin. The study also showed shorter incubation times with the clearing solution resulted in a scaffold with larger ECM fiber diameter that supported higher endothelial cell densities [37]. Histological evidence suggests that decellularized plant tissues are highly biocompatible overall, triggering only a small initial immune response which diminishes over time [24]. To address this issue, mild heat treatments have been explored as an additional step to reduce acute immune response, alter scaffold stiffness, and increase cell viability [42]. Currently, many studies rely on histological evidence and staining procedures to investigate immune responses, so more adoption of alternative methods (i.e., activation studies, protein-based assays, or gene expression assays) may allow for a more robust understanding of how different plant tissue and altered decellularization and functionalization protocols modulate immune responses [43].
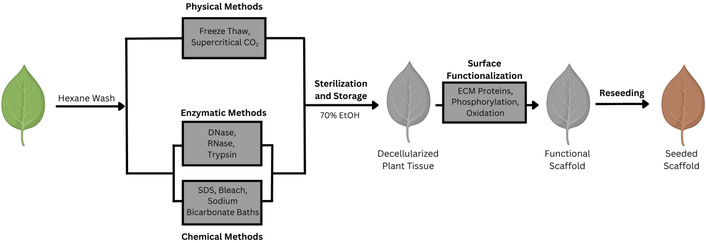
Overview of the decellularization process. A representative decellularization process is depicted based on protocols used in [44] and [37]. Plant tissue can be washed in a hexane bath to remove waxy cuticles before undergoing decellularization by physical, enzymatic, or chemical methods. The resulting decellularized leaves are sterilized and stored in 70% ethanol, functionalized, and then reseeded with fresh cells for the desired engineering structure.
Physical methods for decellularization include the application of freeze-thaw cycles, high hydrostatic pressures, sonication, and supercritical carbon dioxide (scCO2) [45, 46]. Freeze-thaw cycles assist with the decellularization of plant tissues by creating ice-crystals that lyse cellular components while leaving the cell wall relatively intact [45]. ScCO2 possesses properties of both gases and liquids, it’s high density, high diffusivity, and low viscosity, allow it to dissolve efficiently into the liquid phase, easily penetrating plant tissues and causing a reduction in intracellular pH through the formation of carbonic acid which causes lysis of the plant cell membranes while preserving cell-wall architecture and mechanical properties [44]. This is a particularly promising alternative to chemical decellularization methods because of its low toxicity, rapid rate of decellularization, potential ability to both sterilize and decellularize plant tissues simultaneously, and its ability to preserve native plant cell wall architecture [44]. Although chemical decellularization remains the most commonly used method, these benefits may make it a promising method for production of decellularized plant scaffolds at larger scales for clinical applications. Other physical methods for tissue decellularization, such as application of high hydrostatic pressure and sonication have been used to decellularize animal tissues, but they have not been explored in plants [47]. Applications of high hydrostatic pressures (> 600 MPa) physically wash the scaffold with a buffer solution, resulting in the uniform lysis of cell membranes over short incubation times without the use of any chemical agents with the drawback of it being unsuitable to decellularization of more fragile tissues [48]. Sonication uses high-frequency soundwaves to disrupt cell membranes, and is often used in conjunction with chemical detergents [49]. Currently, robust optimizations of these decellularization protocols, which characterize the corresponding changes in mechanical properties, cytocompatibility, immunogenicity, and degradability in response to changing decellularization conditions, are limited to chemical and enzymatic decellularization methods [37]. However, robust optimization of this step to understand the relationship between decellularization and scaffold properties could be highly productive. An optimization study has shown that alterations in these protocols change the mechanical properties and microstructure of the decellularized plant matrix [50]. Therefore, future work could benefit from exhaustive optimization which characterizes how different decellularization protocols alter properties that greatly impact scaffold functionality. This work could improve the reproducibility of standardized scaffolds which will eventually be critical to efforts to introduce these technologies into a clinical setting.
After decellularization, what remains of the plant tissue is primarily the plant cell wall, a specialized type of ECM composed of various polysaccharides, structural proteins, and polyphenolic molecules (see Figure 2) [51]. All plant cells form a malleable primary cell wall comprised mostly of cellulose microfibrils, hemicelluloses, and pectins which allow for the cell’s controlled expansion [52]. The approximate composition of primary cell walls is 15–40% cellulose, 30–50% pectic polysaccharides, and 20–30% hemicellulose (based on cell wall dry weight); however, notable exceptions exist such as grasses, where arabinoxylans transiently make up 20–30% of the primary cell wall, and sugar beet and celery parenchyma which contain high levels of cellulose and pectin but very little hemicellulose [53]. After a plant cell is done growing, secondary cell walls are produced in certain plant cell types to provide significant additional mechanic strength [52]. These cell walls are characterized by large quantities of cellulose (up to 40–50% of cell wall dry weight) and the presence of lignin, a polymer comprised of sinapyl alcohol (creates syringyl, S units), coniferyl alcohol (creates guaiacyl, G units) and p-hydroxylphenyl (H units), which coat cellulose microfibrils and significantly rigidify the cell wall [54, 55]. Cellulose itself is a polymer composed of linear β-1,4 linked glucose molecules. These linear polymers form microfibrils that make up the backbone of the cell wall, and are in part tethered together by structurally diverse hemicelluloses and pectins [56, 57]. Overall, the mechanical properties of plant cell walls can vary considerably between species and tissue types by altering the structural composition or ratios of the polymers in the wall [53]. For example, pectic polysaccharides (e.g., homogalacturonan, rhamnogalacturonan I and II) and hemicelluloses such as xylan, xyloglucan, galactan, mannan, glucan, and glucomannan, possess remarkable structural diversity as a result of complex branching and side-chain modifications [58–60]. Further details on the composition of the plant cell wall, its formation, and the specific branching patterns and composition of hemicelluloses and pectins are reviewed in [52] and [61].
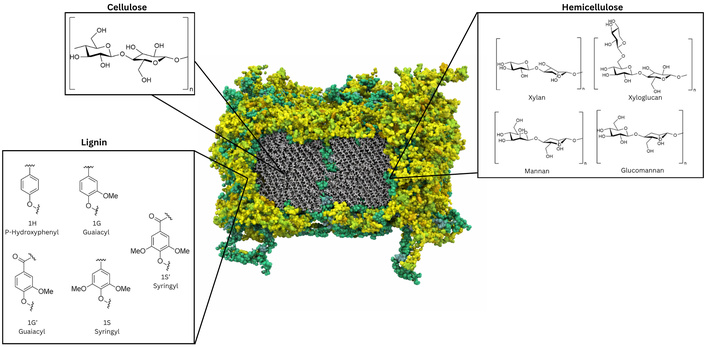
Structure overview and chemical building blocks of the plant cell wall. A 3D macromolecular model of a Populus secondary cell wall adapted from one of our previous publications by Addison et al. [54], © The Author(s) 2024. CC-BY 4.0. Cellulose microfibrils (grey) are surrounded by lignin (yellow) and hemicellulose (green) content. Chemical structures of the backbone of cellulose (β-1,4 linked glucose monomers), the hemicelluloses xylan, xyloglucan, mannan, and glucomannan, as well as the monomeric building blocks of lignin, namely, sinapyl alcohol (syringyl, S units), coniferyl alcohol (guaiacyl, G units), and p-hydroxylphenyl (H units) are shown; their molecular structures were produced using ChemDraw V. 23.1.1.
Notably, altering the composition and mechanical properties of the plant cell wall through genetic engineering has been extensively explored for the purposes of making designer plants as bioenergy feedstocks [62]. For example, in Arabidopsis thaliana, repression of lignin biosynthesis pathway enzymes can reduce lignin content by up to 50% [63, 64]. Cellulose, hemicellulose, and lignin content can be modified similarly through genetic engineering, with up or down regulation of specific enzymes or transcriptional regulators having significant effects on plant cell wall composition, morphology, and ultimately mechanical properties [56, 64]. Since the mechanical properties of plant cell wall are influenced by its composition, which is tunable through genetic engineering, this represents a potential opportunity to tune the mechanical properties of biological tissues while retaining their unique features like inbuilt vascularization.
Due to the accessible hydroxyl groups abundantly present in the polysaccharide components on the surface of the plant cell wall, decellularized plant scaffolds are amenable to chemical modification and functionalization (see Table 1). Although some studies demonstrate that certain cell types can adhere to the decellularized plant scaffold without modification [65], many employ a protein coating (such as fibronectin or collagen) or chemical modifications to improve cell adhesion, alter the material properties of the scaffold (porosity, rigidity, roughness, etc.), or enhance its biodegradation. The earliest plant decellularization work used type I collagen, glutaraldehyde, and fibronectin coatings to enhance scaffold biocompatibility [23, 26], and while these are still used in studies today the breadth of these types of modifications has greatly expanded. Oxidation of the cellulose scaffolds by oxidizing agents such as sodium periodate, sodium bromate, or sodium hypochlorite to convert the 1,2-dihydroxyl groups on cellulose to dialdehydes has been explored to improve the biodegradability of plant-based scaffolds at the cost of some mechanical strength as demonstrated by a reduction of the scaffold’s compressive modulus from 1.52 MPa to 0.6 MPa [32]. Reacting the resultant aldehyde with an organosilane, a silicon-based coupling agent, has shown promise in creating scaffolds for bone tissue regeneration by making a scaffold with improved swelling capacity, porosity, and cell adhesion capacity while retaining its improved degradability [66]. In a Borassus flabellifer endosperm scaffold, modification with the organosilanes 3-aminopropyltriethoxysilane (APTES) and octadecyltrichlorosilane (OTS) increased scaffold degradation from 5% to 16.91% and 13.38% of total scaffold mass respectively over three weeks [66]. Surface modifications with grape seed proanthocyanidin-rich extract have similarly been used in bone tissue engineering, with functionalized scaffolds benefiting from the antioxidant and anti-inflammatory properties of the anthocyanidins in addition to showing enhanced osteogenic outcomes such as greater calcium deposition, mineralized nodule formation, and upregulation of osteogenic markers [67]. Although the specific mechanisms behind which compounds achieved these effects are obscured by this untargeted functionalization approach, its success suggests that there may be a wide breadth of bioactive compounds which could be used to modulate scaffold functionality. Other surface modifications using magnesium oxide nanoparticles, and nano amyloid coated with hydroxyapatite have similarly been investigated as surface coatings to enhance bone tissue engineering [68, 69]. Magnesium oxide nanoparticle coatings have been shown to enhance scaffold stiffness, alkaline phosphatase activity (a marker for osteogenic differentiation), and calcium deposition [69]. This modification has also been shown to reduce inflammatory responses upon scaffold insertion into a rat scab wound while also enhancing rates of wound closure [66]. Additionally, phosphorylation of plant scaffolds has been shown to offer benefits in bone and dental pulp tissue engineering by enhancing osteogenic differentiation and cell proliferation [70, 71]. Finally, specialized peptides have also been used as surface coatings to enhance cell adhesion to scaffolds. The peptide RGD coupled to dopamine (Arg-Gly-Asp-dopamine) was designed to mimic a motif of adhesive domains found in fibronectin and other ECM proteins to allow for enhanced cell adhesion to the cellulose scaffold [28]. Other researchers have cleverly coupled the use of larger co-polymers to prevent non-specific protein adsorption while retaining only their functional peptides of interest [72]. Collectively, these approaches highlight how deliberate modification of the plant scaffold surface enables control over cell–material interactions, degradation behavior, and lineage-specific differentiation outcomes.
Functionalization, mechanical properties, and applications of decellularized plant tissues.
| Plant tissue | Application | Functionalization | Mechanical properties | Tested in vitro/in vivo |
|---|---|---|---|---|
| Apple | Bone tissue engineering [23, 73], probiotic delivery [74], liver tissue engineering [75] | None | YM = 1.1 ± 0.1 kPa [23], APD = ~310 µm [74] | In vivo |
| Bone tissue engineering [23] | Collagen | YM = 2.2 ± 0.2 kPa [23] | In vitro | |
| Bone tissue engineering [23] | Glutaralahyde | YM = 4.1 ± 0.3 kPa [23] | In vitro | |
| Alstroemeria stem | Peripheral nerve healing [76] | None | Average elongation = 7.95%, σmax = 0.02676 N/mm2 | In vivo |
| GelMA | Average elongation = 4.92%, σmax = 0.02204 N/mm2 | In vivo | ||
| Asparagus officinalis | Neural stem cell culture [77] | Poly-L-ornithine | EM = 128 ± 20 kPa, average pore diameter of 39 ± 15 µm | In vitro |
| Bamboo stem | Bone tissue engineering [32] | None | Pore size = 20–150 µm, CS = 1.52 ± 0.346 MPa | In vivo |
| Oxidation (sodium periodate) | CS = 0.6 ± 0.045 MPa | In vivo | ||
| Borassus flabellider endosperm | Bone tissue engineering [66] | None | APD = 19.2 µm, Porosity = 75.84 ± 2.75%, CM = 0.59 ± 0.19 kPa | In vivo |
| APTES (3-aminopropyl-triethoxysilane) | APD = 18.2 µm, Porosity = 79.33 ± 2.05%, CM = 5.73 ± 1.76 kPa | In vivo | ||
| OTS (methyl-terminated octadecyltrichlorosilane) | APD = 7.0 µm, Porosity = 77.96 + 1.27%, CM = 8.33 ± 1.52 KPa | In vivo | ||
| Celery | Skeletal muscle engineering [78] | None | APD = 133 ± 96 μm, Porosity = 70% | In vitro |
| Dill (Anethum graveolens) leaves | Adipose tissue 96modeling [79] | GelMA | YM = 3.55 ± 0.34 kPa, CM = 12.49 ± 1.67 kPa, σmax = 2.19 ± 0.18 kPa, εres = 5.45 ± 0.81% | In vitro |
| Fennel (Foeniculum vulgare) leaves | Adipose tissue modeling [79] | GelMA | YM = 4.27 ± 1.16 kPa, CM = 19.86 ± 3.06 kPa, σmax = 3.28 ± 0.62 kPa, εres = 4.22 ± 0.87% | In vitro |
| Green onion | Skeletal muscle engineering [8] | Fibronectin | Groove size: inner (12 μm × 20–30 μm × 10 μm) and outer (9 μm wide × 10–15 μm) | In vitro |
| Leatherleaf viburnum | Cardiac and vascular tissue engineering [50, 80] | SDS decellularization + glutaraldehyde + fibronectin | APD = 13.9 to 15.4 µm [80] | In vitro |
| None + SDS decellularization | APD = 15.8 ± 2.2 µm [80], EM = 4.0 ± 1.4 [50] | In vitro | ||
| None + SDS/EGTA decellularization | APD = 12.9 ± 1.2 μm [80], EM = 3.9 ± 0.3 MPa [50] | In vitro | ||
| None + tergitol/EGTA decellularization | EM = 2.7 ± 0.9 MPa [50] | In vitro | ||
| None + tergitol/SDS decellularization | EM = 2.8 ± 0.2 MPa [50] | In vitro | ||
| Lotus Petiole | Neural cell alignment monitoring [81] | Graphene oxide | Not characterized | In vitro |
| Lotus stem | Bone tissue engineering [82] | None | Porosity = 88.66 ± 1.76%, CS = 0.49 ± 0.02 MPa, TS = 0.20 ± 0.05 MPa, APD ≈ 130 µm | In vivo |
| Mineralization with CaCl2 and K2HPO4, pH 11 ammonia incubation | Porosity = 67.33 ± 0.88%, CS = 8.54 ± 0.36 MPa, TS = 1.24 ± 0.03 MPa, APD ≈ 130 µm | In vivo | ||
| Mineralization (as above) + PVA hydrogel + manganese carbonyl (MnCO)-loaded mesoporous polydopamine (mPDA) microspheres | Porosity = 64 ± 2.64%, CS = 12.9 ± 0.75 MPa, TS = 1.5 ± 0.1 MPa, APD ≈ 130 µm | In vivo | ||
| Magnolia leaf | Muse cell culture [9] | None | In vitro | |
| Manicaria saccifera palm | Smooth muscle cell, fibroblast, mesenchymal stem cell culture [83] | Alkali treated | YM = 8.22 ± 4.86 GPa, TS = 72.38 ± 45.19 MPa | In vitro |
| Alkali treated + autoclaved | YM = 9.51 ± 4.38 GPa, TS = 68.62 ± 27.93 MPa | In vitro | ||
| Nopal (Opuntia ficus-indica) | Regenerative dentistry; human dental pulp stem cells culture [84] | None | APD = 252 ± 77 μm, TS = 11.8 ± 0.5 MPa | In vitro |
| Onion skin and onion leaf | Vascular patch engineering [85] | Polylactic-co-glycolic acid (PLGA)-based rapamycin nanoparticles | APD = 252 ± 77 μm, TS = 11.8 ± 0.5 MPa | In vivo |
| Parsley stems | Cardiac tissue engineering [86] | None | Burst Pressure = 403.4 ± 83.5 mmHg, EM = 5.182 ± 0.856 MPa, TS = 0.471 ± 0.044 MPa | In vitro |
| Persian walnut (Juglans regia) leaf | Wound dressing [87] | None | TS = 0.60 ± 0.23 MPa, YM = 4.17 ± 0.91 MPa, Porosity = 67.01 ± 1.65% | In vivo |
| Phoenix dactyliferous endocarp | Bone tissue engineering [67] | Grape seed proanthocyanidin extract | APD = ~52 μm, Porosity = ~27% | In vitro |
| Pumpkin | Bone tissue engineering; wound healing [69] | None | APD = 80.96 μm, CS = 0.5633 ± 0.0013 MPa, YM = 3.5 ± 0.05 MPa, TS = 1.42 ± 0.19 MPa | In vivo |
| Magnesium oxide nanoparticles | APD = 70.83 μm, CS = 0.5721 ± 0.0003 MPa, YM = 4.2 ± 0.07 MPa, TS = 1.71 ± 0.31 MPa | In vivo | ||
| Spinach leaf | Cardiovascular tissue engineering [26, 65]; bone tissue engineering [88]; brain vasculature and capillary modeling [89, 90] | None | Maximum Tangent Modulus = 0.3 ± 0.15 [26], APD = 8.7 nm [88] | In vitro |
| Cardiovascular tissue engineering [26, 65] | Fibronectin | Not quantified | In vitro | |
| Cardiovascular tissue engineering [65] | Type IV collagen | Not quantified | In vitro | |
| Sorghum leaves | Skeletal muscle engineering [72] | Poly(PEGMEMA-r-VDM-r-GMA) copolymer + RGD peptide | Not quantified | In vitro |
| Tomato thorny leaf | Tumor microenvironment modeling [91] | Plasma treated with a tabletop plasma cleaner | APD = 6 nm, Total Pore Volume = 0.013 (cm3·g−1) | In vitro |
YM: Young’s Modulus, CM: compressive modulus, EM: Elastic Modulus, TS: Tensile Strength, APD: Average Pore Diameter, σmax: maximum stress, εres: residual strain, GelMA: gelatin methacryloyl.
Decellularized plant tissues have seen significant advancements towards their applications in regenerative medicine for scaffolding the growth of nerve, muscle, cardiac, liver, and bone tissue over the last ten years. Successful efforts often take advantage of similarities in mechanical properties, inherent plant vasculature, or similarities in tissue microstructures between animals and plants. For example, using scanning electron microscopy (SEM) the classic honeycomb-like structure of transverse human skeletal [92] and cardiac muscle [93], porous structure of trabecular bone [94], hexagonal organization of the hepatic acinus [95], and surface structure of decellularized porcine aortas [96] can be roughly observed in celery [78] andgreen onion [8], spinach [89], bamboo [32] and pumpkin [69], and apple [75] respectively. Likewise, each of these decellularized plant scaffolds have been used in successful engineering efforts of its corresponding tissue type. To explore these advances further, this section will briefly review the progress that has been made in tissue engineering for each of these areas.
Cardiovascular disease is still the leading cause of mortality worldwide, so improved methods for healing or replacing damaged cardiac tissue are highly desirable [45]. Implantable tissue-engineered patches incorporating human induced pluripotent stem cell-derived cardiomyocytes (hiPS-CMs) have been proposed as an alternative approach for restoring contractile function to damaged regions of the heart that may otherwise require a transplant [65]. Decellularized spinach leaves have been demonstrated to effectively allow for adherence and growth of hiPS-CMs with or without any additional surface coatings such as human fibronectin and collagen [26, 65]. While these results are promising proof-of-concept for the creation of plant-derived vascular patches, significantly lower contractile strain from hiPS-CMs grown on the spinach leaf scaffolds compared to human myocardium indicates that additional work is needed to understand how contractile strain can be improved to mimic native tissues and ultimately restore native contractile function in vivo [65]. Decellularized parsley stems have also been shown to create a suitable environment for mesenchymal stem cell attachment and growth, allowing for the creation of a pseudo endothelium, and possess suitable mechanical properties making them promising scaffolds for potential tissue-engineered vascular grafts for the purposes of bypassing or replacing blocked vessels [86]. Finally, decellularized leatherleaf viburnum scaffolds have been combined with perfusion bioreactor conditioning to create vascular grafts that demonstrate improved cell viability, longevity, and enhanced endothelial cell density [80].
Significant work has also been carried out to create skeletal muscle tissues using decellularized plant tissue scaffolds. Numerous plants including apple, asparagus, broccoli, carrots, celery, cucumber, green onion, leek, and potato have been tested for their compatibility as scaffolds for skeletal muscle cell growth [8]. Of the scaffolds tested, the white bulb of green onion has been identified to contain an appropriate surface structure to support muscle cell adhesion, differentiation, growth, and myofibril alignment [8]. Decellularized sorghum leaves functionalized with poly(poly(ethylene glycol) methyl ether methacrylate-ran-vinyl dimethyl azlactone-ran-glycidyl methacrylate) (P(PEGMEMA-r-VDM-r-GMA), PVG) coatings were shown to enable the culture of skeletal muscle cells derived from human embryonic stem cell-derived, and their subsequent differentiation to aligned myotubes as well as uniaxial muscle contraction which represents a significant advancement towards creating functional muscle units [72]. Finally, myoblast culture on decellularized spinach leaf scaffolds demonstrated that the intrinsic vascular and fibrous microarchitecture of the plant tissue is sufficient to support cell adhesion, alignment, and the emergence of contractile muscle behavior in vitro, even in the absence of supplemental ECM protein coatings [65].
Considerable progress has been made into bone tissue engineering using decellularized plant scaffolds, with research into this area notably leading to the introduction of new decellularized plant materials and the creation of new surface coatings to enhance scaffold properties. Initial efforts to demonstrate induced osteogenesis in decellularized plant scaffolds evaluated apple, broccoli, carrot, jujube, persimmon, and sweet pepper scaffolds seeded with pluripotent stem cells; however, only decellularized apple scaffolds proved capable of supporting pluripotent stem cell growth [73]. Although these early apple-based engineered tissues lacked a native bony ECM, osteogenically induced cells on decellularized apple scaffolds generated mineralized, bone-like constructs that were successfully implanted into a rat calvarial defect model, demonstrating in vivo biocompatibility, resistance to immune rejection, and the capacity of plant-derived cellulose scaffolds to support functional bone tissue formation [73]. Improved mineralization and osteogenic differentiation, as evidenced by clear overexpression of osteogenic genes, was shortly demonstrated on a decellularized spinach leaf scaffold seeded with mesenchymal stem cells, which are more lineage appropriate for bone tissue engineering than pluripotent stem cells [88]. A key issue with both apple and spinach-based scaffolds was their lack of biodegradability, and thus inability to be replaced with natural host ECMs, due to mammals lacking the necessary enzymes to degrade the polysaccharide components of the plant cell wall. Work with decellularized bamboo scaffolds aiming to take advantage of its bone-like structure [29] attempted to overcome this issue by applying a chemical oxidation approach using a sodium periodate treatment [32]. Although this treatment reduced the scaffold’s mechanical strength, making the scaffolds unsuitable for load-bearing applications, it improved mesenchymal stem cell adhesion and biodegradation as evidenced by a 70% reduction in scaffold size after subcutaneous implantation into a rat for four weeks compared to 40% for a non-modified scaffold [32]. Complementary efforts have focused on imparting bioactivity through biomimetic surface functionalization, such as the incorporation of nano-amyloid fibrils and hydroxyapatite, which together enhanced cell adhesion, osteogenic differentiation, and in vivo trabecular bone formation [68]. A different approach using magnesium oxide nanoparticles enhanced the hydrophilicity, mechanical stiffness, surface roughness, and biodegradation rates of a decellularized pumpkin scaffold, resulting in enhanced osteogenic behavior in adipose-derived mesenchymal stem cells and improved wound closure with suppressed inflammatory responses in rats with incision wounds [69]. Cumulatively, decellularized plant tissues show considerable promise in bone-tissue-engineering applications, having already demonstrated enhanced wound healing, efficient osteogenic differentiation, and early bone formation; however, despite advancements with chemical modifications to scaffold surface structure their immediate clinical applications remain limited by their biodegradability and load-bearing capabilities.
Although still a budding application, decellularized plant tissues have seen some use as novel methods to enhance wound healing and as routes for drug delivery. Vascular patches created from decellularized red onion soaked in polylactic-co-glycolic acid (PLGA)-derived rapamycin nanoparticles have been explored as a potential biomaterial for small diameter (< 6 mm) vascular grafts in place of traditional synthetic materials such as polyester and polytetrafluoroethylene (PTFE) due to their risk of causing neointimal hyperplasia, where vascular smooth muscle cells adhere to the scaffold vasculature causing blockages [85]. In a proof of concept, the decellularized plant tissue has been shown to reduce neointimal thickness while implanted in rat inferior vena cava. Further work into this area achieved even more promising results by combining the PLGA-rapamycin nanoparticles with interleukin-33 (IL-33) antibodies, which bind IL-33 involved in neointimal hyperplasia formation, absorbed into the plant scaffold, resulting in a thinner neointima [97]. Decellularized walnut leaves have been explored as potential biomaterials for improved wound dressings, having been demonstrated to retain some beneficial native bioactive components after decellularization that possess antimicrobial, anti-inflammatory, and antifungal properties while supporting accelerated wound closure [87]. Finally, recently developed decellularized alstroemeria scaffolds have been introduced as the first plant-based nerve conduit effectively applied to improve axonal regeneration in rat models with a 1-cm sciatic nerve defect [76]. These scaffolds were engineered to control the release of insulin-like growth factor-1 LR3 using a niosome-based delivery system to enhance axonal regeneration, and they were further treated with gelatin methacrylate (gelMA) based on a previous report that gelMA hydrogels enhanced Schwann cell adhesion, viability, and maturation [98]. This final example in particular highlights the power of mature decellularized plant scaffold technology, showing how combinations of modifications to the surface chemistry and intelligently designed drug release systems can be used to achieve incredible healing outcomes. Overall, these examples demonstrate the promising potential of using decellularized plant scaffolds in wound healing and drug delivery.
Beyond strictly medical applications, decellularized plant tissues have been investigated for their use as model systems to study biological phenomena without needing live animal models, similar to the application of organoids to study specific disease states [99]. Inspired by the similar diameter (10–300 µm) and shared branching structure of spinach leaf vasculature and human arterioles and capillaries, decellularized spinach leaves have been examined as a platform for studying the impact of magnetic nanoparticles and the thrombolytic drug urokinase on thrombosis induced by creating a synthetic clot composed of platelet-free lyophilized human plasma and thrombin [89]. This approach demonstrated that plant vasculature can serve as functional frameworks for the study of drug delivery and vascular biology studies in vitro. The vasculature of decellularized spinach leaves was further exploited to create a model system for studying vascular amyloidosis, a condition where amyloid fibrils are deposited along blood vessel walls [90]. While this system yielded promising results into its potential for studying the accumulation and clearance of amyloid, technical challenges with preparing thin sections of the scaffold rendered it incompatible with electron microscopy and only mainly compatible with fluorescence microscopy to monitor amyloid deposition and transport within the model.
Decellularized plant scaffolds have also been used to explore how physical context alters cellular responses to therapies. Cells cultured on decellularized spinach leaf scaffolds exhibited significantly different mechano-regulation, morphology, proliferation, and responses to chemotherapeutic drugs and ionizing radiation compared with cells on rigid tissue culture polystyrene [100]. For instance, melanoma and prostate cancer cells seeded on leaf scaffolds showed downregulated YAP/TAZ mechanotransduction signaling, altered morphology, reduced proliferation, and differential drug and radiation responses, reflecting the softer, more tissue-like mechanical environment provided by plant cellulose compared to stiff plastic. This suggests that plant scaffolds can serve as physiologically relevant models for evaluating therapeutic responses and stress signaling that may be obscured in conventional 2D systems.
Beyond vascular and mechanobiological models, plant-derived scaffolds have facilitated adipose and metabolic tissue modeling. Decellularized dill and fennel leaf matrices, when embedded within gelMA hydrogels, formed channel networks that enhanced nutrient transport and supported adipogenic differentiation and lipid accumulation over extended culture, offering a platform for studying adipose tissue physiology and metabolic responses in a 3D context that incorporates both structural and transport cues [79].
The tumor microenvironment has also been recapitulated using plant scaffolds. Decellularized tomato thorny leaves served as a natural 3D scaffold for hepatocellular carcinoma cells, enabling cancer cell proliferation, morphological heterogeneity, and drug resistance patterns that more reflective of in vivo tumors than traditional monolayer cultures [91]. This reflects how scaffold architecture influences phenotypic behaviors relevant to cancer progression and therapeutic screening.
Neural tissue modeling has benefited from plant cellulose scaffolds as well. In early work on neural stem cell culture, decellularized cellulose matrices supported 3D proliferation and differentiation of neural progenitors into neurons and glia with structural complexity absent from 2D culture, underscoring the ability of plant scaffolds to provide mechanical and spatial cues relevant to neural development [77].
Finally, plant scaffolds have recently been applied to reproductive tissue analogues. For example, decellularized celery stalk, green onion leaf, and bluegrass leaf functionalized with type I collagen have enabled the culture of smooth muscle cells in configurations that mimic aspects of the myometrium, providing a model to investigate contractility and hormone responsiveness in vitro [101]. Collectively, these efforts demonstrate the potential low-cost and ethically uncontroversial decellularized plant cell scaffolds offer in tissue and disease modeling.
Current literature is largely in vitro; regulators will expect stepwise in vivo evidence (small to large animal) under good laboratory practice (GLP) to assess local/systemic immune response, in situ degradation/remodeling, mechanical performance under physiological load, and host integration/vascularization over clinically relevant durations [36, 46]. To translate lab-scale decellularization to manufacturing, processes must also be adapted for controlled, reproducible production. Scalability depends on managing plant-source variability and implementing robust unit operations (e.g., perfusion decellularization and controlled oxidation/conjugation) that preserve architecture at scale [46].
Sterilization process needs to be compatible with plantderived cellulose matrices and functional coatings. For preparing decellularized plant tissues, a standard BSL-1 laboratory with good aseptic technique and a Class II biosafety cabinet is generally sufficient, since plant tissues and common decellularization reagents are not associated with human pathogens. For clinical translation, the manufacturing environment is driven by aseptic processing and cleanroom controls rather than high biosafety levels. When terminal sterilization is not feasible (e.g., to preserve bioactive coatings), production should be performed under a validated aseptic process in qualified cleanrooms, in line with related guidelines [102–104]. For reproducibility, ensuring removal of residual chemicals/endotoxins and establishing sustainable processing with stringent quality control (QC) are essential to ensure batchtobatch consistency [36, 46].
Long-term in vivo performance is affected by several factors. First, anatomical mismatch and functional constraints: exactly matching plant microarchitecture to target tissue requirements (pore size, mechanical properties, vascular accessibility, permeability) is challenging [36, 46, 105]. Secondly, mechanical performance and stability: decellularization can change stiffness and strength; many plant scaffolds exhibit limited mechanical robustness, variability between samples, and culture-induced disintegration, constraining use in load-bearing or long-term applications [36, 46]. Thirdly, degradability and persistence: Plant cellulose is slow/poorly degradable in vivo, which can hinder applications where controlled resorption is required; strategies to modulate degradation are needed [36, 46]. For long-term in vivo safety, chronic in vivo studies are needed to assess hemocompatibility and systemic toxicity, where applicable.
Taken together, the advancements discussed here demonstrate that decellularized plant tissues are a highly promising biomaterial for tissue engineering and regenerative medicine. This article presents essential concepts, including key decellularization techniques and scaffold surface modification approaches, while highlighting major developments towards applying decellularized plant scaffolds in and regenerative medicine and tissue engineering. Significant progress has been made in terms of understanding and standardizing the decellularization process with new surface modifications to scaffolds offering exciting routes to fine-tune scaffold performance towards specific applications such as bone tissue engineering. Coupling these understandings of scaffold modifications with their capacity for controlled drug release has so far proven extremely productive as demonstrated with the development of scaffolds capable of enhancing axonal regeneration [76]. Utilization of decellularized plant tissues has expanded beyond their initial scope of just serving as scaffolds for cell proliferation and the plant tissues have emerged as versatile platforms for drug and probiotic delivery [74], alternative approaches to lab-grown meat production [78, 106, 107], and systems for modeling disease states. The ability to readily genetically engineer plant cell wall composition further distinguishes plant-based scaffolds from animal-derived counterparts, potentially offering a unique route to rationally design scaffold mechanics and architecture at the source of plant cells.
However, despite these immense successes, challenges remain. Certain applications, such as kidney tubule engineering, have failed when attempted with chive and spinach leaves due to anatomical restrictions preventing the spreading of cells throughout the scaffold vasculature, insufficient permeability, and rapid solute diffusion [105]. Many studies evaluating a wide range of scaffolds have found that many are unsuitable for their intended applications. For example, early bone tissue engineering efforts tested apple, broccoli, sweet pepper, carrot, and jujube, but they only found decellularized apple to support viable culture of pluripotent stem cells [73]. A more comprehensive and standardized understanding of the available decellularized plant tissues and functionalization methods could make it easier to narrow down a suitable scaffold for a desired engineering goal, making previously inaccessible engineering goals possible without extensive testing of scaffolds. Additionally, while there has been significant progress in terms of standardizing chemical and enzymatic decellularization approaches [36], significant experimental space remains unexplored. An extensive statistical design of experiments, with more diverse decellularization reagents, different plant tissues, and precise measures of success such as the immunogenicity of the scaffold, mechanical properties, degradation and cell seeding efficiency, could lead to a more robust understanding of which decellularization techniques are most efficient for a desired application.
Looking forward, the continued improvement of decellularized plant scaffold technology will likely depend on integrating advanced surface chemistries, dynamic remodeling capabilities, and controlled delivery of bioactive factors with scalable and reproducible manufacturing processes. In parallel, expanding their use as physiologically relevant in vitro models may accelerate discoveries in drug screening, mechanobiology, and disease modeling while reducing reliance on animal systems. Together, these developments position decellularized plant tissues as a distinct and versatile biomaterial platform with the potential to reshape regenerative medicine and experimental biology.
ECM: extracellular matrix
GelMA: gelatin methacrylate
HiPS-CMs: human induced pluripotent stem cell-derived cardiomyocytes
IL-33: interleukin-33
PLGA: polylactic-co-glycolic acid
scCO2: supercritical carbon dioxide
SDS: sodium dodecyl sulfate
The views expressed in the article do not necessarily represent the views of the DOE or the U.S. Government. The U.S. Government retains and the publisher, by accepting the article for publication, acknowledges that the U.S. Government retains a nonexclusive, paid-up, irrevocable, worldwide license to publish or reproduce the published form of this work, or allow others to do so, for U.S. Government purposes.
AL: Conceptualization, Writing—original draft, Writing—review & editing. HW: Conceptualization, Writing—review & editing. YJB: Writing—review & editing, Supervision. All authors read and approved the submitted version.
The authors declare that they have no conflicts of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
This material is based upon work at the Center for Bioenergy Innovation supported by the U.S. Department of Energy, Office of Science, Biological and Environmental Research under Contract Number [ERKP886]. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 754
Download: 21
Times Cited: 0
Aylin Kara Özenler ... Funda Tihminlioglu
Damla Arslantunali Sahin ... Vasif Hasirci
Momoko Ebisawa ... Yutaka Inoue
Athira S. Dev ... Renu Mohan
Viktor Borysiuk ... Serhii Nedilko
