 Review
Review

Affiliation:
1Department of Biotechnology and Genetic Engineering, Jahangirnagar University, Dhaka 1342, Bangladesh
Email: taslimsajib@gmail.com
ORCID: https://orcid.org/0009-0002-1696-8158
Affiliation:
2British Columbia School, Dhaka 1207, Bangladesh
ORCID: https://orcid.org/0009-0006-6514-9860
Affiliation:
3Department of Medicine, Sir Salimullah Medical College & Mitford Hospital, Dhaka 1100, Bangladesh
ORCID: https://orcid.org/0009-0006-0141-1866
Affiliation:
4Chittagong Medical College, Chittagong 4203, Bangladesh
ORCID: https://orcid.org/0009-0005-9675-4257
Affiliation:
5Department of Public Health and Informatics, Bangladesh University of Professionals, Dhaka 1216, Bangladesh
ORCID: https://orcid.org/0009-0005-2586-6980
Affiliation:
3Department of Medicine, Sir Salimullah Medical College & Mitford Hospital, Dhaka 1100, Bangladesh
ORCID: https://orcid.org/0009-0005-6594-1789
Affiliation:
6Department of Educational and Counselling Psychology, University of Dhaka, Dhaka 1000, Bangladesh
ORCID: https://orcid.org/0009-0000-4197-1522
Affiliation:
7Barind Medical College and Hospital, Rajshahi 6207, Bangladesh
ORCID: https://orcid.org/0009-0002-3086-1189
Affiliation:
8Rajshahi Medical College, Rajshahi 6000, Bangladesh
ORCID: https://orcid.org/0009-0004-0361-4726
Affiliation:
9Department of Surgery, Dhaka Medical College Hospital, Dhaka 1000, Bangladesh
ORCID: https://orcid.org/0009-0007-0291-0584
Affiliation:
10MRes Public Health, University of Wolverhampton, Wolverhampton WV1 1LY, UK
ORCID: https://orcid.org/0009-0009-6136-4100
Affiliation:
11Khulna City Medical College Hospital, Khulna 9000, Bangladesh
ORCID: https://orcid.org/0009-0006-0776-5382
Affiliation:
12Shaheed Ziaur Rahman Medical College, Bogura 5800, Bangladesh
ORCID: https://orcid.org/0009-0000-1596-6295
Affiliation:
13Govt. Pioneer Women’s College, Khulna 9100, Bangladesh
ORCID: https://orcid.org/0009-0007-7953-5137
Affiliation:
14Department of Zoology, Ain Shams University, Cairo 11566, Egypt
ORCID: https://orcid.org/0009-0007-3733-5890
Explor Neuroprot Ther. 2026;6:1004155 DOI: https://doi.org/10.37349/ent.2026.1004155
Received: December 17, 2025 Accepted: April 22, 2026 Published: May 25, 2026
Academic Editor: Abdelhamid Benazzouz, Bordeaux University, France; Rafael Franco, Universidad de Barcelona, Spain
Norepinephrine (NE), a central catecholamine neurotransmitter synthesized primarily in the locus coeruleus (LC), plays a critical role in regulating arousal, attention, emotional processing, and stress responsiveness. While contemporary personality neuroscience has established the role of NE in acute psychological states, its contribution to stable personality traits remains underexplored. This review synthesizes neurobiological, psychological, genetic, and psychopharmacological evidence to propose a NE-personality continuum that links tonic and phasic dynamics of the LC-NE system to enduring individual differences in alertness, anxiety, and adaptability. Alertness is associated with optimal noradrenergic tone and efficient phasic signaling, which enhances the signal-to-noise ratio and attentional focus. Anxiety arises from chronic hyperactivation or dysregulated NE release, particularly involving excessive α1- and β-adrenergic receptor activity and impaired modulation from the prefrontal cortex. Adaptability denotes a harmonious interaction between the limbic system and prefrontal cortex, which facilitates cognitive flexibility and emotional regulation in response to changing environmental demands. The connection between NE activity and personality traits follows an inverted U-shaped pattern. Low tone leads to apathy and less engagement, moderate tone helps with resilience and optimal functioning, and high tone leads to hypervigilance and rigidity. This model combines findings from fundamental neuroscience and clinical research to provide a physiologically based framework for understanding how long-term variations in noradrenergic regulation affect personality traits, as described in established trait theories. The findings underline the feasibility of adding noradrenergic biomarkers and pharmaceutical therapies into clinical practice, as well as the importance of longitudinal and multimodal research to determine trait-level causality. This is especially important for understanding how to use these elements to improve treatment plans for personality disorders.
Norepinephrine (NE) is a central catecholamine neurotransmitter that modulates arousal and information processing, thereby influencing behavioral and cognitive responses to environmental demands [1, 2]. Personality is the enduring pattern of thinking, feeling, and behaving that defines individual differences [3]. Personality traits are most commonly conceptualized within the Big Five personality traits framework, which includes openness, conscientiousness, extraversion, agreeableness, and neuroticism. Within this structure, anxiety-related tendencies align with neuroticism, alertness overlaps with conscientiousness and aspects of extraversion, and adaptability relates to openness and emotional stability, reflecting cognitive flexibility and regulatory balance [4, 5]. “Alertness” is defined as a neurocognitive state of arousal and vigilance, not a personality trait, in this review. However, individuals show that baseline arousal regulation and responsiveness can be varied, suggesting that alertness could be a trait-like feature throughout time and situations. Alertness, not a personality trait, is studied at the junction of state-dependent cognitive performance and biologically limited personality diversity [6].
Temperament models such as Cloninger’s Temperament and Character Theory further emphasize biologically grounded dispositional systems, linking harm avoidance to anxiety, novelty seeking to exploratory engagement, and persistence to sustained attentional control. Similarly, Jeffrey Alan Gray’s Reinforcement Sensitivity Theory associates anxiety with heightened behavioral inhibition system (BIS) activity, involving noradrenergic mechanisms [7, 8]. Although these models acknowledge monoaminergic influences, contemporary personality neuroscience has under-integrated tonic and phasic noradrenergic dynamics. This review addresses that gap by mapping alertness, anxiety, and adaptability onto established trait frameworks and proposing a NE-personality continuum grounded in locus coeruleus -NE (LC-NE) regulation. Conceptually, alertness overlaps with conscientiousness and aspects of extraversion in the Big Five personality traits, anxiety corresponds to neuroticism and harm avoidance in temperament models, and adaptability relates to openness and emotional stability. Thus, the proposed NE-personality continuum does not introduce new traits but offers a neurobiological explanation for variation within established personality dimensions.
Beyond classical trait models, some neurobiologically grounded frameworks have attempted to map personality onto specific neuronal systems. Many personality models provide background for the current framework; for example, Colin DeYoung’s Cybernetic Big Five Theory links personality traits to large neural networks that emphasize goal-oriented information [9]. Another personality model (dopaminergic), proposed by Richard Depue, focuses on reward sensitivity and motivation in trait and behavioral approaches. Jaak Panksepp identified evolutionarily conserved emotional circuits that underpin personality from an affective neuroscience perspective, whereas behavioral inhibition or activation systems (BIS/behavioral activation system) are biologically based personality traits in Jeffrey Gray’s paradigm. Although these advances have been made, nothing has been done to study how NE affects stable individual differences in arousal regulation and alertness control [10].
The LC is the primary site of synthesis for NE, sometimes referred to as noradrenaline, which is broadly distributed throughout the brain, limbic system, and spinal cord [11, 12]. The noradrenergic system contributes to the complex construction of personality by serving as a crucial neuromodulatory framework that regulates behavioral and emotional inclinations [13, 14]. Its effect can be viewed as a continuum in which individual differences in tonic activity and responsiveness lead to various personality traits [15]. The system’s reactivity is another important component of neuroticism; a hyperresponsive noradrenergic system lowers the stress threshold, leading to a propensity for anxiety and negative emotionality [16, 17]. This same overactivity can also manifest as heightened aggression and emotional sensitivity due to a generalized state of hypervigilance and a reduced capacity to control emotions. Consequently, an individual’s unique patterns of involvement, emotional reactivity, and adaptation to the social and stressful demands of external stimuli are shaped by a stable neurobiological substrate provided by the noradrenergic system [18, 19]. However, low NE levels caused by long-term stress, neurodegenerative illnesses such as Parkinson’s disease, and neurodevelopmental disorders such as attention-deficit/hyperactivity disorder (ADHD) can result in sadness, exhaustion, and apathy, and a reduction in alertness, focus, and concentration. In this context, NE functions as a natural stimulant that enhances focus and alertness. When released from the LC, it sharpens the mind, heightens alertness, and amplifies the senses, all of which help organisms recognize, evaluate, and react to threats more skillfully [20, 21]. Furthermore, disorders such as post-traumatic stress disorder (PTSD), generalised anxiety disorder, and panic disorder can exhibit symptoms of anxiety, hyperarousal, irritability, and hypervigilance owing to malfunctioning noradrenergic signaling, which is caused by high NE levels or dysregulated release from the LC [22]. However, by improving the signal-to-noise ratio in various brain regions, such as the prefrontal cortex and the sensory cortex, NE helps to improve cognitive focus and attention by suppressing distractions and improving the processing of pertinent and noteworthy stimuli [23]. When the level of NE is ideal, it enhances behavioral flexibility by modifying working memory and attention in the prefrontal cortex, which is crucial for adjusting to a changing environment [24]. Though there is a lot of evidence that NE is linked to transient mental states, its role in forming stable personality traits is not yet well understood in modern personality neuro-based research frameworks. Both dopaminergic and serotonergic mechanisms are focused on by existing models, neglecting the role of tonic and phasic LC-NE dynamics in persistent individual variations in alertness control and regulation [25].
The purpose of this review is to clarify how the noradrenergic system plays a significant physiologic role in personality. It looks at how its actions, from consistent control to fast reactions, affect important characteristics such as stability and indifference, as well as anxiety and rigidity. Additionally, this review looks at how the noradrenergic system affects three interconnected personality-related dimensions that are crucial for individual diversity and survival: attentiveness, anxiety, and adaptability. This review investigated how individual differences in alertness, anxiety, and adaptability are influenced by variations in NE activity. This paper offers an updated view of how the noradrenergic system contributes to the complex architecture of personality traits by combining findings from neurobiological, psychological, and clinical studies. There is still a lack of knowledge regarding how individual variations in noradrenergic activity affect long-lasting personality traits, despite compelling evidence connecting NE to psychological states, including alertness and anxiety [26]. Current models do not connect NE levels to personality-related dimensions; instead, they focus on short-term pharmacological effects or clinical symptoms [27]. To create a NE-personality continuum and show how different types of noradrenergic regulation contribute to characteristics such as alertness, emotional stability, and adaptability that define typical behavior patterns, this review attempts to integrate neurobiological, genetic, and psychopharmacological evidence. This is a narrative review, theory-building study aimed at synthesizing current neurobiological and psychological evidence to construct a conceptual framework linking NE dynamics to personality traits. Relevant literature was identified through a structured, non-systematic search of major scientific databases, prioritizing recent and high-impact studies. A thematic synthesis approach was employed to integrate findings across experimental, clinical, and theoretical research.
NE is synthesized from dopamine via the enzyme dopamine β-hydroxylase and is released in response to both internal and external stimuli [28, 29]. Dopamine β-hydroxylase makes NE from dopamine, and NE is released when the body or the outside world sends signals. The LC-NE system is the main source of central NE and is very important for controlling arousal, attention, stress response, and cognitive control [30, 31]. This system modulates brain states that produce stable differences in how people react to emotions and how much attention they pay to things by sending signals to many brain regions, including the cortex and limbic system [32, 33]. NE works on three main types of adrenergic receptors: α1, α2, and β. Each type has its own set of functions. α1 receptors are involved in sympathetic arousal, which causes vasoconstriction, pupil dilation, and increased alertness during the fight-or-flight response [34, 35]. When central α1 receptors are too active under stress, they can make the prefrontal cortex work less well and make anxiety worse. Inhibitory feedback mechanisms regulate NE release via presynaptic α2 autoreceptors. Despite a comprehensive understanding of the receptor-level processes, their connection to personality remains ambiguous [36]. It can be seen from current research that these regulatory systems primarily affect transient alertness control rather than persistent trait variations, suggesting that while they influence short-term states of arousal, they may not significantly impact long-term personality traits [37]. Peripherally, β receptors speed up the heart rate (HR), increase the amount of blood pumped by the heart, and open up the airways. Centrally, they help consolidate emotional memories, especially in the amygdala. So, whether NE makes you more alert (through balanced α2A signaling) or more anxious and hyperaroused (through too much α1 and β activation) depends on which receptor subtype is activated [38, 39].
Both tonic and phasic types of activation are present in the LC-NE system. Interpreting noradrenergic activity requires distinguishing transient state effects from enduring trait characteristics. Most experimental research manipulates NE acutely (e.g., pharmacological challenges) to measure arousal and attention [40]. However, personality qualities need consistency over time and context. There is little longitudinal evidence that baseline LC-NE dynamics predict long-term personality characteristics [41]. Tonic exercise promotes wakefulness and baseline alertness, both of which are essential for cognitive performance. The maximum performance and alertness periods of sustained and concentrated concentrations are associated with moderate tonic LC activity. The tonic alertness that supports other cognitive processes is provided by this intermediate level of activity, which is linked to good task performance [42, 43]. Phasic bursts are associated with adaptive responses and task-related concentration. NE release at particular cortical and subcortical destinations increases quickly during phasic bursts, increasing neuronal gain for task-relevant stimuli. By maximizing the signal-to-noise ratio in sensory and associational circuits, this mode makes it easier to identify noteworthy events and permits a quick, precise behavioral response. Thus, stable characteristics associated with attention control, affective regulation, and behavioral flexibility, fundamental aspects of personality, may be supported by individual variations in LC-NE function [44]. Variations in LC-NE activity are believed to influence consistent behavioral patterns, particularly in areas such as attention regulation and stress response. Differences in baseline pupil-linked arousal and LC functional connectivity have been associated with variations in attentional performance and cognitive control [45]. However, most of this information comes from cross-sectional or task-oriented studies, which limit the ability to draw conclusions about long-term personality stability. As a result, the current findings should be interpreted as reflective of trait-like tendencies rather than stable personality traits themselves [46]. Table 1 shows both tonic and phasic modes of activity with distinctive characteristics in the LC-NE system.
Locus coeruleus-norepinephrine (LC-NE) system illustrating both tonic and phasic modes of activity.
| Feature | Tonic mode | Phasic mode | References |
|---|---|---|---|
| Pattern | A slow, sustained, regular discharge of single spikes | A brief, high-frequency discharge with a rapid & high spiking activity | [47] |
| Frequency | 0.1–5.0 Hz | 10–20 Hz | [48] |
| Trigger | General behavioral state and arousal | Elicited by novel or salient polymodal sensory stimuli | [49, 50] |
| Function | Maintenance & Regulation of the general state of arousal, vigilance, and alertness | Facilitates the significance & relevance of an event and orients precise responses to allocate and prioritize resources to relevant information | [48] |
| Effect | Establishes the background level of norepinephrine release | Creates high frequency and sharp spikes in NE release | [51] |
Long-lasting individual differences in personality may be explained neurophysiologically by the balance of tonic and phasic NE release. Because their system is suitably primed for phasic reactions to stimuli, people with steady, moderate tonic baselines are likely to be emotionally resilient and adaptive. A low tonic state, on the other hand, results in apathy and low levels of engagement since the system is unable to mobilize energy for motivated action, whereas a person with a chronically high tonic state develops a personality marked by neuroticism and hypervigilance. It is hard to measure central NE directly in people, so indirect proxies are often used. Pupillometry is a non-invasive indicator of LC activity, whereas a functional neuroimaging tool (e.g., functional magnetic resonance imaging [fMRI]) evaluates LC activation and its connectivity with prefrontal and limbic areas [52–54]. Pharmacological agents like clonidine and propranolol elucidate noradrenergic influences on behavior. Peripheral metrics such as HR variability (HRV) and galvanic skin response (GSR) are utilized as indicators of sympathetic activation. These methods enable the investigation of the relationship between LC-NE responsivity and trait-level variations in alertness, anxiety, and adaptability [55, 56].
Multiple approaches index LC-NE activity, but each presents significant drawbacks that hinder interpretation. For example, pupillometry serves as a non-invasive proxy for LC activity and offers high temporal sensitivity at the cost of neurochemical specificity; however, pupil size is influenced by various neuromodulatory systems, including cholinergic and dopaminergic inputs, which limit its specificity for NE [57]. fMRI of the LC enhances spatial localization, but sacrifices signal reliability and temporal precision due to the LC’s small size and its proximity to cerebrospinal fluid (CSF) [58]. Additionally, HRV and GSR are indirect indicators of sympathetic activation that fail to distinguish between central noradrenergic activity and other autonomic functions. Consequently, these measurements reflect LC-NE function probabilistically rather than definitively [59].
Although these effects are often characterized by overall noradrenergic tone, this approach does not fully address the complexities at the receptor level. Noradrenergic tone by itself cannot account for the functional effects of NE, as the activities of specific receptor subtypes introduce complications [60, 61]. In particular, α2-adrenergic receptor subtypes exhibit diverse and sometimes contradictory functions. A decline in NE tone within the prefrontal cortex results in diminished activation of postsynaptic α2A receptors, adversely affecting working memory and concentration while contributing to fatigue. Conversely, heightened activity of α2C receptors, especially during synaptic NE release or with pharmaceutical agonists, is linked to lethargy, sedation, and diminished arousal. Pharmacological data indicate that α2 agonists such as clonidine induce more sedation than guanfacine, possibly due to variations in selectivity for the α2C versus α2A receptors [62]. Therefore, inadequate NE signaling and receptor-specific inhibitory mechanisms may lead to fatigue and reduced alertness, highlighting the receptor-dependent and non-linear nature of the noradrenergic system [63].
In summary, the mechanistic neurobiology of NE is well understood; however, its relevance to personality neuroscience remains unclear. Although the connections between tonic and phasic LC activity and trait-like differences are intriguing, they currently lack empirical support. Challenges such as measurement specificity, reliance on indirect proxies, and cross-sectional study designs complicate causal inference. Critically, the predominance of cross-sectional and pharmacological challenge designs limits causal inference, making it unclear whether LC-NE dynamics reflect stable personality-related dimensions or transient state-dependent processes [64]. Future research that is longitudinal, genetic, and multimodal will help clarify whether LC-NE dynamics play a role in enduring personality traits or primarily reflect adaptive, state-dependent processes [65].
Importantly, alertness is treated here as a state-dependent neurocognitive construct with trait-like variability, rather than a discrete personality trait. NE has both short-term effects that depend on the state and longer-term effects that depend on the trait. Acute variations in LC-NE activity influence transient arousal, vigilance, and stress responses, indicative of state changes [66]. Neurocognitive dimensions, on the other hand, are characterized by consistent behavioral patterns over time and across different situations. Consequently, experimental manipulations of NE predominantly reveal state effects, whereas trait-level interpretations necessitate evidence of consistent interindividual variations in tonic or phasic LC-NE regulation [67]. The capacity to maintain focus and be prepared to react to changes in the surroundings is known as alertness. By improving the signal-to-noise ratio in cortical processing, NE increases attentional focus and sensory sensitivity [23, 68, 69]. High NE levels are correlated with vigilant personality-related dimensions characterized by quick responsiveness and heightened situational awareness [70]. On the other hand, low-energy, inattentive, or lethargic traits, such as some depressive or apathetic traits, have been linked to decreased NE activity [71, 72]. Insufficient NE release from the LC is associated with fatigue, diminished concentration, and reduced motivational drive [73]. When noradrenergic tone is too low, cortical signal amplification is weakened, leading to impaired attentional focus and decreased responsiveness to environmental demands. Such hypoactivation contributes to lethargy, inattentiveness, and limited behavioral adaptability, features commonly observed in depressive or apathetic states [74]. LC phasic activation is responsible for this effect. It selectively increases evoked responses to salient stimuli while suppressing spontaneous brain activity, resulting in more accurate sensory representations and increased attentional concentration [75]. The “adaptive gain” theory of the LC-NE function, which holds that optimal phasic activity promotes behavioral flexibility and better performance in activities demanding alert attention and quick reactions to environmental changes, explains this correlation. Noradrenergic arousal, which ranges from low to ideal to high, has a direct effect on personality by converting chemical states into stable traits like resilience, anxiety, or indifference [48].
NE levels and cognitive function have an inverse U-shaped relationship. Reduced awareness and trouble focusing are signs of inadequate cognitive performance at low levels. Important processes such as focused attention, behavioral flexibility, and general cognitive performance are improved when NE levels reach a modest, ideal range. However, when the level of NE is too high, it causes a reduction in cognitive control, which manifests as increased anxiety, stiff thinking, and difficulty in focus because of hypervigilance [18, 76–78]. Individual diversity in the LC response is correlated with trait-like variations in alertness, indicating a biological foundation for alertness and sustained attention.
Furthermore, a biological connection between noradrenergic tone and individuals with higher baseline alertness is supported by research employing psychophysiological measures such as pupil dilation, a surrogate for LC-NE activation, which reveals that those with higher baseline NE reactivity exhibit improved attentional performance [79, 80].
Figure 1 below illustrates how alertness, defined as the ability to sustain attention and respond rapidly to environmental changes, is regulated by NE activity originating from the LC. At the top, the figure presents an inverted-U curve showing the relationship between NE levels and cognitive performance. Low NE levels are associated with reduced alertness, lethargy, and inattentive personality traits. As the NE increases to an optimal range, the sensory sensitivity, signal-to-noise ratio, and attentional focus improve, resulting in enhanced cognitive function and behavioral flexibility. Excessively high NE levels are linked to anxiety, hypervigilance, cognitive rigidity, and impaired focus.
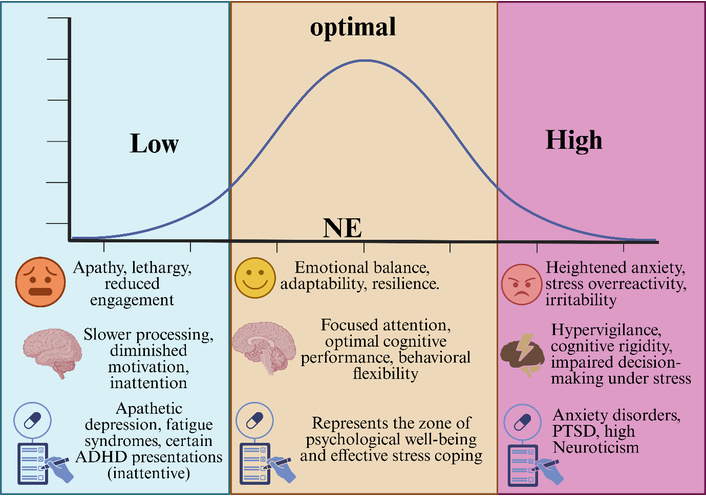
Noradrenergic modulation of alertness and cognitive performance. ADHD: attention-deficit/hyperactivity disorder; NE: norepinephrine; PTSD: post-traumatic stress disorder.
While converging evidence associates NE activity with alertness and attentional performance, numerous studies depend on acute pharmacological interventions or indirect indicators like pupillometry, constraining inferences regarding stable trait variations. Most findings are cross-sectional, and few longitudinal studies show that baseline NE responsivity is a good predictor of persistent personality-related dimensions. More precise methods and forward-looking designs are necessary to elucidate trait-like effects [81]. Taken together, the evidence suggests a consistent association between NE activity and attentional processes; however, the lack of longitudinal and trait-focused designs prevents definitive conclusions regarding stable personality differences [82].
Excessive activation of NE causes anxiety and hyperarousal, but it also encourages adaptive attentiveness. Vigilance and threat sensitivity are increased by increased NE activity, particularly in the prefrontal cortex and amygdala [83, 84]. Exaggerated phasic responses and tonic hyperactivity are hallmarks of excessive and dysregulated NE discharge from the LC, which is crucial in interfering with activity in the amygdala and prefrontal cortex [85]. Excess NE activity in the amygdala enhances fear responses, which are major factors in the manifestation of pathological anxiety and hyperarousal in stress-related disorders such as depression and PTSD in the form of core symptoms such as sleep disturbances, irritability, and heightened startle responses. Despite the considerable evidence indicating that increased noradrenergic activity enhances amygdala reactivity and anxiety-like behavior, most of these studies rely on neuroimaging or pharmacological challenge paradigms involving small sample sizes. This is largely due to the fact that much of the research focuses on pharmacological modifications or the induction of acute stress, which complicates the ability to draw definitive conclusions about stable trait anxiety. Consequently, these limitations make it challenging to extrapolate the findings to real-world scenarios, where anxiety can manifest in various forms [86–88]. When there is a lot of stress, too much stimulation of α1-adrenergic receptors in the prefrontal cortex makes executive control worse by weakening working memory networks and encouraging bottom-up limbic dominance. More α1 activation changes how the brain processes information from reflective to reflexive responses. This makes people more alert, causes emotional problems, and makes anxiety symptoms worse [89]. Individuals with high trait anxiety often exhibit overactive noradrenergic signaling, leading to exaggerated stress responses and difficulty in regulating emotional states [90–93]. Moreover, excessive NE release results in heightened physiological arousal and hypervigilance. Elevated NE levels amplify amygdala reactivity and sympathetic activation, increasing susceptibility to anxiety, panic responses, and exaggerated threat perception. Under these conditions, excessive arousal disrupts prefrontal regulation, impairing cognitive flexibility and reducing the ability to adapt appropriately to changing environmental cues [94].
Importantly, anxiety regulation is a dynamic interaction between the sympathetic noradrenergic system and the parasympathetic cholinergic system. NE activates the LC to cause sympathetic arousal, while acetylcholine (ACh) does the opposite through the vagus nerve to improve parasympathetic tone and promote physiological calm [95, 96]. Higher vagal tone is linked to better control of emotions, less activity in the amygdala, and better control of the prefrontal cortex. So, anxiety can come from both too much NE signaling and not enough parasympathetic counter-regulation. Therapeutic techniques that enhance vagal tone, such as slow breathing, biofeedback, mindfulness, and vagus nerve stimulation, might reestablish autonomic equilibrium by fortifying cholinergic regulation of stress responses [97–101]. This connection is supported by pharmacological evidence: Palpitations and tremors are among the physical signs of anxiety that are lessened by β-adrenergic blockers, which decrease NE signaling. Propranolol is a beta-blocker that is used to treat high blood pressure (BP), tachycardia, and anxiety symptoms such as palpitations, sweating, and trembling. Propranolol is a nonselective beta-adrenergic antagonist that competes with catecholamines such as adrenaline and noradrenaline for beta-1 and beta-2 receptors in the sympathetic nervous system. Blocking these receptors efficiently lowers the peripheral physiological sensations of stress, thereby altering the brain feedback loops that might exacerbate anxiety [102, 103].
However, moderate NE activity is beneficial, as it sharpens perception and promotes adaptive concern in uncertain situations [104–106]. Moderate NE levels increase emotional regulation by increasing the ability of the brain to concentrate, process information, and manage stress responses. Moderate NE promotes prefrontal modulation of limbic pathways, allowing balanced emotional processing. This finding is consistent with results that neither hypoactivity nor hyperactivity promotes adequate emotional stability [107]. Thus, the connection between NE and anxiety exhibits an inverted U pattern, with both under- and overactivation impairing normal emotional control [108, 109]. A significant portion of the evidence linking NE to anxiety originates from clinical populations, specifically PTSD/ADHD, in which dysregulation may not represent typical personality variation [110]. The results from clinical populations reveal significant dysregulation in extremes of noradrenergic function. However, these findings may not be applicable to normative personality variation, where the effects are likely to be more nuanced and context-dependent [111]. There should be concern while applying these results to normal trait anxiety. Moreover, discrepancies in neuroimaging and pharmacological studies indicate that NE probably interacts with other systems, highlighting the necessity for integrative and longitudinal research [110].
Adaptability, the ability to adapt behavior in changing environments, is dependent on flexible cognitive and emotional regulation. The LC-NE system adjusts attention and decision- making in response to novelty and uncertainty [112–114]. Adaptive individuals show balanced NE reactivity, allowing them to remain alert yet not overwhelmed by stress [115, 116]. Conversely, dysregulated NE signaling is associated with rigid or maladaptive features, which are shown in diseases such as ADHD and PTSD, where people find it difficult to correctly change their focus or emotion [117]. Coordinated activity between the LC, prefrontal cortex, and anterior cingulate cortex regions influenced by NE is necessary for maximum adaptation, according to functional imaging studies. This neurochemical equilibrium promotes psychological resilience by allowing people to alternate between exploratory and goal-directed behaviors [48, 118]. The brain’s capacity to adjust to changes throughout life is known as neural flexibility, or neuroplasticity, and it manifests as observable personality traits that define an individual [119]. Neural flexibility plays the function of exploratory plasticity in terms of an individual’s openness to experience, where a propensity for novel stimuli forces the brain to form new, expansive neural connections, leading to sensory awareness and intellectual curiosity [120, 121]. Flexibility plays a role in regulatory stability in conscientiousness. Consistent goal-directed behavior creates top-down control networks in the prefrontal cortex, which strengthen effective neural connections and improve planning and impulse control [122–124]. Therefore, the basic biological basis for these unique, observable behavioral patterns is individual variations in the baseline capability for neuroplastic changes. People with effective LC-prefrontal cortex (PFC) coupling typically exhibit characteristics linked to resilience, openness, and cognitive flexibility [125, 126].
Theoretical models suggest that flexible LC-PFC coupling facilitates adaptability; however, empirical evidence is predominantly indirect and cross-sectional. Only a limited number of studies directly examine whether individual variations in NE dynamics forecast long-term personal stability. Subsequent research employing longitudinal, genetic, and multimodal methodologies will be crucial for determining causal and persistent associations [60, 127]. This review discusses noradrenergic pathways; however, it emphasizes that personality-related processes arise from interactions within neuromodulatory systems rather than solely from neurotransmitters. Dopaminergic pathways influence reward sensitivity, motivation, and exploratory behavior, which are fundamental to extraversion and cognitive flexibility. Serotonergic systems play a role in regulating moods, behavior, and emotions, interacting dynamically with NE [128]. The modulation of prefrontal cortex function by dopamine can either mitigate or enhance the effects of noradrenergic influences on attentional control, while serotonergic regulation of limbic activity helps to limit hyperarousal. The NE-personality continuum should be understood within a broader monoaminergic network framework. This perspective helps to avoid simplistic interpretations and provides a clearer representation of the decentralized neurochemical basis of personality [129].
The role of NE in personality can be conceptualized as a continuum of arousal regulation [130].
Low NE tone: A low NE tone is associated with fatigue, diminished motivation, and reduced alertness, leading to slower cognitive processing and decreased engagement with the environment [108, 125, 131, 132].
Moderate NE tone: A moderate NE tone supports focused attention, emotional balance, and adaptability, enabling optimal cognitive performance and flexible responses to changing demands [107, 133, 134].
High NE tone: A high NE tone is linked to hypervigilance, heightened anxiety, and increased rigidity, often resulting in overreactivity to stress and impaired decision-making under pressure [18, 135].
It is essential to interpret this continuum within the context of receptor subtype-specific effects (see the Neurobiology section), as similar behavioral outcomes, such as fatigue or reduced alertness, can be caused by both low NE levels and increased α2C-mediated inhibitory signaling [136]. To synthesize these data, the NE-personality continuum develops an integrative model that places personality qualities along a noradrenergic tone continuum. Fatigue and low motivation observed in certain clinical presentations are caused by low NE tone at one endpoint of the continuum. An ideal zone for attention, emotional stability, and adaptability that defines resilience is created by a moderate NE tone. High NE tone causes hypervigilance, anxiety, and cognitive rigidity, which are essential components of neuroticism at the other end of the scale. While this continuum is supported by converging experimental and clinical findings, direct empirical validation at the trait level remains limited, and the model should be interpreted as a heuristic framework rather than a confirmed biological taxonomy. By showing how stable individual differences in NE regulation from genetic risk factors to medication response create the constellation of traits that characterize an individual’s distinctive patterns of behavior, cognition, and emotion, as discussed in Table 2 below, this model synthesizes the evidence [18].
Effects of low, moderate, and high norepinephrine (NE) tone on personality traits and their different contributions.
| NE tone | Characteristics | Impact on personality | References |
|---|---|---|---|
| High | Anxiety, restlessness, hypervigilance, and insomnia | Results in elevated alertness, anxiety, sleep disturbances, and hyperactivity | [137, 138] |
| Moderate | Balanced energy, optimal focus, alertness & arousal, and stable mood | Optimal cognitive function, emotional stability & motivation | [70] |
| Low | Lethargy, fatigue, brain fog, inability to concentrate, and impaired alertness | Lack of motivation, Anhedonia, apathy, emotional blunting | [139] |
Additionally, this continuum combines personality traits with tonic/phasic LC patterns, indicating that interindividual variance in alertness, emotional stability, and adaptability is biologically based on stable differences in NE dynamics.
Understanding the noradrenergic basis of personality may have potential implications for clinical and personality research [140]. The identification of positive emotional cues was improved in a clinical setting by pharmacologically produced elevations in noradrenaline levels. In-group bias and out-group disparagement are associated with noradrenergic transmission, indicating their influence on intergroup relationships and social identity. Pharmacological modulation of NE (e.g., with reuptake inhibitors or β-blockers) may influence personality-related affective states [141, 142]. For example, personality-related affective states, especially high-arousal negative emotions, are directly impacted by pharmaceutical manipulation of NE with β-blockers such as propranolol [143]. While β-adrenergic receptors play a role in the peripheral symptoms of anxiety, such as tachycardia and tremors, anxiety disorders are not solely caused by the activation of β-receptors. Central mechanisms involving α1- and α2-adrenergic receptors, along with interactions with limbic and prefrontal networks, significantly influence the cognitive and affective dimensions of anxiety [144]. Thus, β-blockers like propranolol effectively mitigate somatic symptoms of performance anxiety; however, they are not regarded as first-line treatments for generalized anxiety disorders, for which selective serotonin reuptake inhibitors (SSRIs) and cognitive behavioral therapy (CBT) exhibit greater efficacy [145, 146]. The capacity of propranolol to cross the blood-brain barrier enables it to modulate central adrenergic activity; however, its main therapeutic indications are hypertension and angina [147]. Research shows that beta-adrenergic blockade with propranolol efficiently attenuates unpleasant, high-arousal emotions such as anxiety in response to psychosocial stresses; this fundamental action is crucial to its effect on affective features. This happens in tandem with a reduction in sympathetic nervous system reactivity, demonstrating a clear mechanism by which noradrenergic intervention can moderate the strong emotional reactivity linked to characteristics such as neuroticism [148]. Psychotherapy and stress management can indirectly normalize NE activity through regulation of the autonomic nervous system [149, 150]. The reformation of maladaptive thinking patterns and the development of adaptive coping mechanisms are facilitated by psychological interventions such as CBT [151].
Although the main effect of drugs is to suppress emotional reactions, relying too much on them to manage stress may ignore the potential advantages of nonpharmacological treatments, which frequently produce better long-term outcomes. A promising future is provided by personalized medicine, which adapts treatments to each patient’s particular genetic composition and way of life [152, 153]. Healthcare professionals can develop individualized stress management programs that maximize treatment efficacy and reduce potential side effects by taking into account a patient’s unique biological characteristics. These plans may involve a mix of medication, therapy, and lifestyle modifications [154–156]. Clinical evaluation, psychological theory, common-sense theories of etiology, and useful stress-reduction techniques are often integrated into psychological treatments. In psychology, stress-reduction techniques often support healthy coping methods [157]. By predicting susceptibility to anxiety or stress-related diseases, NE responsivity may eventually contribute to personalized therapy, although current evidence remains insufficient for clinical implementation [158]. Individual differences in noradrenergic reactivity can be assessed using both central and peripheral biomarkers. Peripheral indicators include HR, BP, HRV, and GSR (electrodermal activity), which reflect sympathetic activation. Reduced HRV, in particular, is often interpreted as diminished vagal tone and heightened sympathetic dominance [159, 160]. Pupil dilation is increasingly used as a non-invasive proxy of LC activity. In research settings, plasma or CSF NE levels, pharmacological challenge paradigms (e.g., clonidine suppression tests), and functional neuroimaging of LC activation provide more direct assessments of central noradrenergic responsivity [161]. When exposed to chronic stress, an individual is more likely to experience symptoms of anxiety and depression if their LC-NE system is more responsive [71, 115].
Numerous pathways linked to the emergence of anxiety disorders are impacted by central noradrenergic signaling. Defensive response and fear conditioning entail arousal, memory formation, consolidation, and retrieval [162]. The functions of the motor cortex, medial prefrontal cortex/anterior cingulate cortex, anterior insula, amygdala, hippocampus, and thalamus have all been connected to these regions. Therefore, an intermediate phenotype of altered central noradrenergic signaling during fear processing and sympathetic noradrenergic activation may be represented by the fine-tuning of these neuronal systems of the defensive motivational system by the noradrenergic system. During fear processing and anxiety, central and peripheral sympathetic noradrenergic activation is controlled by a MIR579 genetic variant that fine-tunes noradrenaline homeostasis [163–165]. Furthermore, there are important translational ramifications for comprehending individual NE responsiveness. Stronger fear acquisition, poor extinction learning, and an increased incidence of anxiety disorders, PTSD, and stress-related psychopathology have all been linked to elevated NE reactivity [94]. Clinically, NE responsivity may be used to predict who is more susceptible to long-term hyperarousal; who might benefit from NE-modulating medications such as prazosin, propranolol, or serotonin-norepinephrine reuptake inhibitors (SNRIs); and who might react better to behavioral interventions such as exposure therapy, which depends on the ideal levels of NE for extinction learning. Therefore, combining genetic, physiological, and neuroimaging markers of NE function may offer a precision-medicine method for detecting high-risk patients and customizing treatment plans [166–170]. Candidate biomarkers like pupillometry, HRV, and neuromelanin-sensitive MRI offer indirect indicators of LC-NE function, yet their specificity is constrained. These measures are affected by various neurochemical systems and contextual elements, reducing their accuracy as independent indicators of noradrenergic tone [171]. Furthermore, variability among laboratories and task paradigms hinders reproducibility. At present, NE-related biomarkers should be considered as probabilistic indicators rather than conclusive diagnostic tools [172].
It is noticeable that noradrenergic biomarkers and pharmacological modulation are effective in some ways but not yet standard in clinical accuracy. Common biomarkers such as pupillometry (the measurement of pupil size and reactivity), HRV, and fMRI lack specificity and consistency for therapeutic applications. The majority of evidence comes from controlled laboratory or clinical settings, with little real-world or longitudinal validation. NE-based profiling to predict personality traits or guide individualized therapies has yet to be developed. While translational applications of NE-based profiling are conceptually promising, clinical implementation encounters practical challenges, including cost, technical expertise, and standardization across settings. Additionally, variations in neuromodulatory systems are contingent upon context, which constrains the viability of consistent trait-based stratification. Therefore, future advancements will hinge on the combination of multimodal biomarkers with longitudinal validation to ascertain clinical utility [173–175]. It is important to note that much of the available evidence remains indirect, frequently cross-sectional in design, and often derived from clinical or high-risk populations. As such, causal inferences and generalization to broader populations should be made with caution [176].
This review provides a thorough perspective on personality by identifying NE as a vital neurobiological regulator that influences long-term individual differences in alertness, anxiety, and adaptability. NE affects attentional focus, emotional reactivity, stress responses, and behavioral flexibility through both tonic and phasic activity patterns in the LC-NE system. The proposed NE-personality continuum illustrates how personality traits can be viewed as a spectrum of noradrenergic tone. Low NE activity correlates with tiredness, lack of motivation, and reduced mental engagement. In contrast, moderate NE levels foster optimal alertness, emotional stability, and adaptability. Excessive NE activity, however, is associated with hypervigilance, anxiety, and cognitive rigidity. Importantly, the connection between NE and personality is not a straight line and depends on the situation. It looks like an inverted U-shaped function, where both hypoactivation and hyperactivation make psychological functioning worse. A major limitation of the current literature is the absence of longitudinal, multimodal studies directly linking LC-NE dynamics to stable personality traits, which contain the ability to distinguish trait effects from adaptive state responses. Although converging evidence substantiates the involvement of noradrenergic mechanisms in the development of trait-like behavioral tendencies, existing findings predominantly consist of cross-sectional data, frequently sourced from clinical populations. Subsequent investigations utilizing longitudinal, genetic, neuroimaging, and multimodal biomarker methodologies are imperative to elucidate causal pathways and trait stability. In a clinical context, comprehending individual variations in NE responsivity may offer potential directions for future tailored interventions, although further validation is required, encompassing pharmacological modulation, psychotherapeutic approaches, and stress-regulation methods aimed at achieving autonomic equilibrium. Noradrenergic activity functions within extensive monoaminergic networks, necessitating that comprehensive models incorporate interactions with dopaminergic and serotonergic systems. In conclusion, the NE-personality continuum proposes a biologically informed conceptual framework that connects neurochemical regulation to enduring personality-related dimensions. This model successfully amalgamates neurobiology with personality theory, augmenting our comprehension of human behavioral diversity in a coherent and practical manner.
ADHD: attention-deficit/hyperactivity disorder
BIS: behavioral inhibition system
BP: blood pressure
CBT: cognitive behavioral therapy
CSF: cerebrospinal fluid
fMRI: functional magnetic resonance imaging
GSR: galvanic skin response
HR: heart rate
HRV: heart rate variability
LC-NE: locus coeruleus-norepinephrine
NE: norepinephrine
PFC: prefrontal cortex
PTSD: post-traumatic stress disorder
TU: Conceptualization, Supervision. OHB: Methodology, Formal analysis. MMM: Supervision, Writing—original draft. SS: Investigation, Writing—original draft. FS: Resources, Writing—review & editing. ST: Project administration, Validation. SA: Writing—review & editing. RS: Formal analysis, Resources, Supervision. RSN: Supervision, Data curation. KYG: Supervision, Writing—original draft. MTH: Project administration. ATA: Supervision. TTI: Writing—review & editing. JF: Writing—original draft. FHA: Investigation, Methodology. All authors read and approved the submitted version.
The authors declare no conflicts of interest.
Not applicable.
Not applicable.
Not applicable.
Not applicable.
This research received no specific grant from any funding agency.
© The Author(s) 2026.
Open Exploration maintains a neutral stance on jurisdictional claims in published institutional affiliations and maps. All opinions expressed in this article are the personal views of the author(s) and do not represent the stance of the editorial team or the publisher.
Copyright: © The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.

View: 4167
Download: 32
Times Cited: 0
