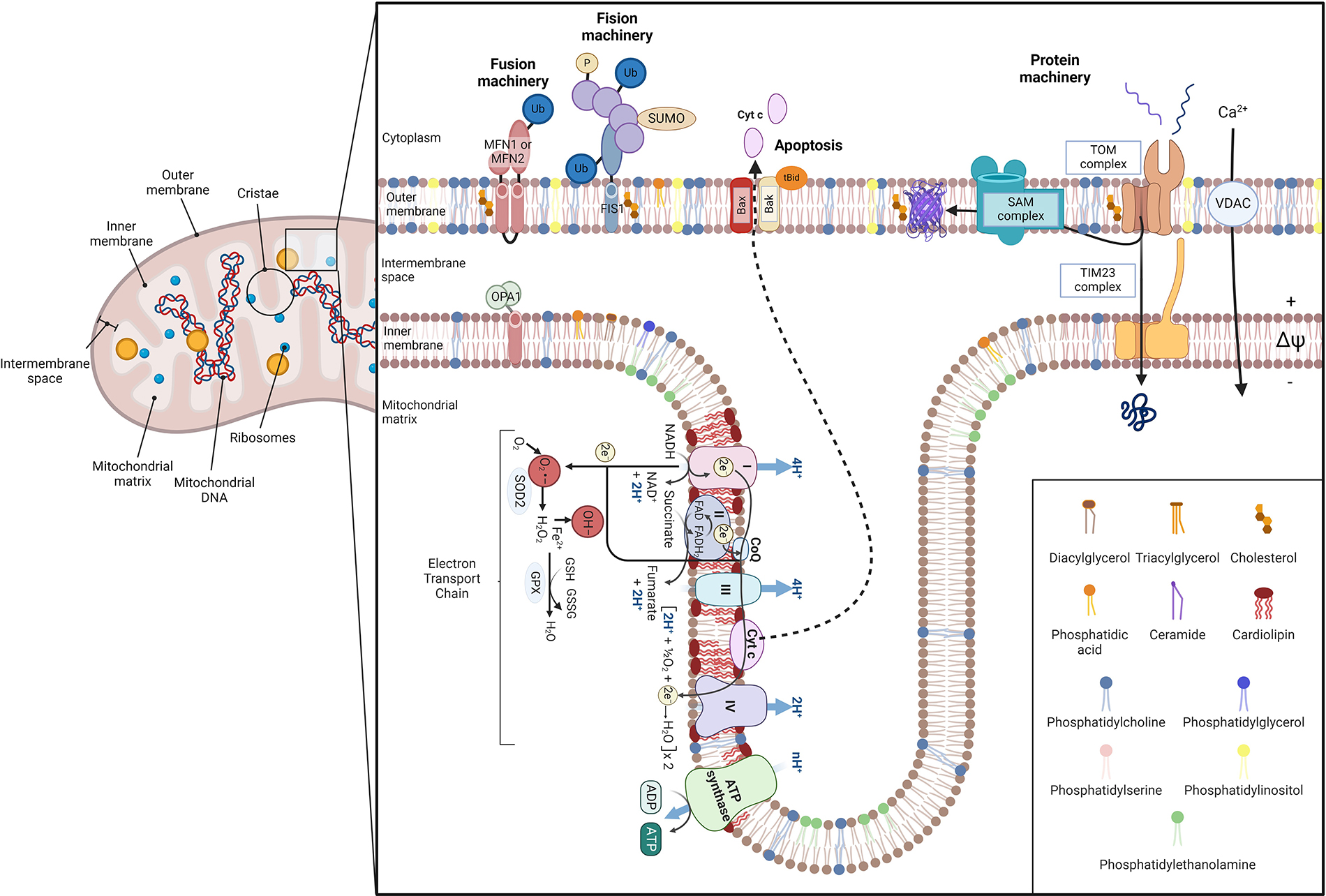
Schematic illustration of mitochondrial architecture and function. Mitochondria are conformed by two bilayers separated by the intermembrane space: the outer mitochondrial membrane (OMM) and the inner mitochondrial membrane (IMM) [1–3]. The IMM surrounds the mitochondrial matrix, where mitochondrial DNA (mtDNA) and ribosomes are located [4], and it includes the inner boundary membrane (IBM) and mitochondrial cristae. Each bilayer has a specific lipid composition and IMM is specially enriched in proteins [7]. Concretely, mitochondrial cristae contain the respiratory chain complexes and the F1F0-ATP synthase, harboring oxidative phosphorylation to obtain energy. The overflow of electrons between each complex generates reactive oxygen species (ROS) that are reduced by mitochondrial antioxidants, such as glutathione (GSH) [8]. ROS can impair mitochondrial constituents, such as mtDNA, phospholipids, and proteins, leading to cytochrome c (Cyt c) detachment from IMM and releasing it into the cytosol via Bax/Bak oligomerization [4, 5, 9]. Furthermore, ROS production can be enhanced by an altered calcium (Ca2+) homeostasis. Mitochondrial dynamics depend on fusion/fission processes. Fusion relies on mitofusin 1/2 (MFN1/2) of the OMM, and optic atrophy protein 1 (OPA1), placed at IMM. In contrast, fission is regulated by dynamin-related protein 1 (Drp-1), which interacts with fission protein 1 (FIS1) [10, 11]. Mitochondria also have a protein machinery to translocate proteins into the matrix. This machinery is composed of the translocase of the outer membrane (TOM) channel the sorting and assembly machinery (SAM) in the OMM and the translocase of the inner membrane (TIM23) in the IBM [12–17]. Ub: ubiquitin; SUMO: small ubiquitin-like modifier; tBID: proapoptotic truncated BID; VDAC: voltage-dependent anion channel; SOD2: superoxide dismutase 2; GPX: glutathione peroxidase; GSSG: glutathione disulfide; O2•−: superoxide; OH–: hydroxyl radical
Note. Adapted from “Electron Transport Chain”, “Mitochondrial Membrane (Phospholipid Bilayers)”, and “Protein Import into the Mitochondria”, by BioRender.com (2024). Retrieved from https://app.biorender.com/biorender-templates